Abstract
Introduction
Hypoxylaceae (Xylariales, Sordariomycetes) is a diverse family of mainly saprotrophic ascomycetous fungi, commonly found in angiosperm-dominated forests. Members of Hypoxylaceae have a worldwide distribution and are characterized by diverse lifestyles, including saprotrophic, endophytic, and phytopathogenic modes of life, colonizing wood, leaves, and fruits, while some species are symbiotically associated with insect vectors in both terrestrial and aquatic habitats (Ju & Rogers 1996; Pažoutová et al. 2013; Wibberg et al. 2021; Cedeño-Sanchez et al. 2024a, b). They are not only associated with woody substrates but have also been recovered from seed plants, lichens, and algae and have even been found in insect nests and sponges (Pažoutová et al. 2013; Medina et al. 2016; U’Ren et al. 2016; Leman-Loubière et al. 2017; Sir et al. 2019). The family shows remarkable diversity in morphology and secondary metabolite production, characterized by the presence of stromatal pigments and a nodulisporium-like anamorph (Wendt et al. 2018; Wibberg et al. 2021).
Historically, taxonomic delineation within Hypoxylaceae relied heavily on morphological characteristics (Ju & Rogers 1996). However, this approach proved insufficient due to phenotypic plasticity and convergent evolution, leading to misclassifications and unresolved species complexes, such as the Hypoxylon fuscum and H. rubiginosum groups (Lambert et al. 2021). With the advent of molecular phylogenetic studies based on ITS, LSU, rpb2, and tub2 dataset, together with chemotaxonomy as an additional tool have significantly improved the resolution of taxa within the family. This has enabled the segregation of new genera such as Hypomontagnella, Jackrogersella, and Parahypoxylon (Lambert et al. 2019; Cedeño-Sanchez et al. 2023). Polyphasic taxonomy, which integrates morphological, chemotaxonomic, and molecular data, has become the standard for Hypoxylaceae classification. Despite these advances, challenges persist. Phylogenetically, certain genera remain polyphyletic, and intragenomic polymorphisms in ribosomal DNA (rDNA) loci, particularly the ITS region, show high intragenomic polymorphism and limited resolution at the species level (Stadler et al. 2020; Cedeño‑Sanchez et al. 2023). Additionally, the cryptic diversity observed in stromatal pigments and secondary metabolite profiles underscores the limitations of relying solely on these markers. For example, many specimens classified as H. rubiginosum (often reported as H. cf. rubiginosum) show molecular differences but lack diagnostic morphological and chemical differences (Pourmoghaddam et al. 2020). These limitations highlight the need for alternative genetic markers. Recent research demonstrates that mitogenomes can provide robust, lineage-specific markers. For instance, in Ganoderma, the atp9 and nad4L genes have proven to be reliable molecular markers and potential DNA barcodes, offering valuable insights into the phylogeny of the genus and the Ganodermataceae family (Chen et al. 2024).
Despite their potential, mitogenomic data remain scarce, with fewer than 0.02% of described fungal species sequenced (Fonseca et al. 2021), representing a critical gap that hinders the resolution of taxonomically complex groups such as Hypoxylaceae. The primary value of this study lies in contributing complete mitogenome sequences, which provide a foundational resource for future comparative genomics. Expanding mitogenomic datasets is essential for future research aiming to elucidate fungal evolution, diversity, and classification in greater detail. Given that only about 0.02% of available data currently contribute to resolving these aspects, broader and more representative taxonomic sampling is urgently needed. Increasing both taxonomic breadth and depth of mitogenomic sampling will allow researchers to evaluate mitogenome utility across hierarchical levels and to integrate mitochondrial markers with existing polyphasic frameworks for more robust species delimitation and phylogenetic inference. This will enable a thorough assessment of the usefulness of mitogenomic data for studies on fungal classification, diversity, and phylogeny, as this field remains in its preliminary stages.
This study presents an extensive analysis of the mitochondrial genomes of 50 Hypoxylaceae specimens, offering a comprehensive dataset that will serve as a foundation for future evolutionary and taxonomic research. By combining traditional methods (morphology and phylogeny) with mitochondrial genome data, 16 novel Hypoxylaceae species are introduced, and insights into their mitochondrial genome structure are provided. Further in-depth analysis and comparison of these mitochondrial genome data will be presented in subsequent articles.
Materials and methods
Sample collection
The fungal specimens were collected during surveys conducted in the tropical and subtropical regions of China from 2019 to 2021. All relevant habitat information was recorded in detail. Specimens were photographed using a Canon G15 camera (Canon Inc., Tokyo, Japan), placed in paper bags, and transported to the laboratory for further examination. To remove excess humidity, they were dried using a portable fan dryer. The dried specimens were carefully labelled and stored in an ultra-low freezer at -80 °C for one week after isolation to eliminate any insects and their eggs. Afterwards, the specimens were subjected to morphological analysis. All specimens were deposited at the herbarium of Guizhou Medical University (GMB) and the Herbarium of Cryptogams, Kunming Institute of Botany, Chinese Academy of Sciences (KUN-HKAS). Living cultures were deposited at the Guizhou Medical University Culture Collection (GMBC). All scientific names of fungi follow the entries in MycoBank and Index Fungorum, hence no authorities and years of publications are given in the text apart from the taxonomic entries. Colors and codes were determined using the color chart of Rayner (1970).
Isolation and morphological characterization
Macroscopic characteristics were examined under an Olympus SZ61 stereomicroscope (Japan) and photographed with a Canon 700D digital camera (Canon Inc., Tokyo, Japan). The samples were mounted in water for microscopic examination, and Melzer's reagent was added to assess the amyloid reaction of the ascal apical apparatus. The Tarosoft (R) Image Framework (v 0.9.7) program and Adobe Photoshop CS6 software (Adobe Systems, USA) were used for measuring and processing images. Axenic cultures were obtained from single spores or tissues as described by Senanayake et al. (2020). Germinating spores were observed with a Stereo Zoom microscope and transferred to potato dextrose agar (PDA; 39 g/l distilled water, Difco potato dextrose). The cultures were incubated at 25 °C for 4–6 weeks, with weekly observations of growth and development.
Mycelial fermentation and DNA extraction
Pure colonies were transferred onto PDA medium and incubated at 25°C for 7 days. Under sterile conditions, a sterile cork borer was used to create holes (0.3 cm in diameter) in the colonies, and 10 mycelial plugs were transferred into Erlenmeyer flasks containing 100 mL of PD liquid medium (Potato Dextrose Liquid Medium). The flasks were then incubated in a rotary shaking incubator at 25°C and 120 rpm for 10 days. After fermentation, the mycelium was filtered out. The filtered mycelium was subjected to DNA extraction using the commercially available BIOMIGA Fungal Genomic DNA Extraction Kit (Biomiga, San Diego, USA). Extracted DNA was used for Sanger sequencing of target loci (ITS, LSU, tub2, rpb2) and whole-genome sequencing (WGS).
Sanger Sequencing for Multi-Locus Gene Regions
Internal transcribed spacers (ITS), large subunit (LSU), tub2 (β-tubulin), and rpb2 (RNA polymerase II subunit) regions were PCR-amplified using primer pairs ITS1/ITS4 (White et al. 1990; Gardes & Bruns 1993), LR0R/LR5 (Vilgalys & Hester 1990), Bt2a/Bt2b or T1/T22 (Glass & Donaldson 1995; O'Donnell & Cigelnik 1997), and rpb2-5F/rpb2-7cR (Liu et al. 1999), respectively. The 25 μL PCR mix included 9.5 μL ddH2O, 12.5 μL PCR Master Mix, 1 μL of each primer, and 1 μL template DNA. Amplification followed the thermal cycling conditions outlined in Samarakoon et al. (2022). PCR products were verified via 1.5% agarose gel electrophoresis with GoldenView staining. Successful amplicons were sent to Sangon Biotech (Sangon Co., China) for sequencing.
Raw sequence chromatograms were manually inspected and quality-trimmed using Geneious Prime V2023.2.1. Forward and reverse reads for each locus were assembled into consensus sequences. The final sequences for each locus (ITS, LSU, tub2, rpb2) were deposited in the National Center for Biotechnology Information (NCBI) GenBank database (Supplementary Table 9).
Whole-genome sequencing (WGS), assembly and annotation
Genomic DNA was sent to Sangon Biotech Co., Ltd. (Shanghai, China) for next-generation whole-genome sequencing and library preparation. The sequences obtained were proofread and then assembled using the software Geneious Prime V2023.2.1. The raw sequencing data underwent initial quality control using FastQC v0.11.9. Adapters and low-quality bases were trimmed using Trimmomatic v0.39. The following thresholds were applied: LEADING:3 TRAILING:3 SLIDINGWINDOW:4:15 MINLEN:50. The mitochondrial genome was assembled from the cleaned reads using SOAPdenovo2 v2.04 with a range of k-mer size and other parameters set to default. Open reading frames (ORFs) were identified using Geneious tools (Kearse et al. 2012). Annotation of the genomes was performed with MFannot (https://megasun.bch.umontreal.ca/apps/mfannot/) (Valach et al. 2014) and MITOS web server (http://mitos.bioinf.uni-leipzig.de/) (Bernt et al. 2013), adhering to the mitochondrial genetic code 4. Manual adjustments were made to the annotations, with reference to the mitochondria of closely related species within the same family. Finally, graphical maps of the Hypoxylaceae mitogenomes were drawn using OGDraw (https://proksee.ca/projects/new) (Grant et al. 2023). The tRNA genes in the 50 Hypoxylaceae mitogenomes were also predicted using tRNAscan-SE v1.3.1 (Lowe & Chan 2016). Assemblies have been deposited in GenBase and GenBank accessions mentioned in Supplementary Table 9.
Mitogenome sequence analysis
The base composition of the 50 Hypoxylaceae mitogenomes was analyzed using the DNASTAR Lasergene v7.1 (http://www.dnastar.com/). Strand asymmetry of the 50 mitogenomes was assessed according to the following formulas: AT skew = (A – T) / (A + T), and GC skew = (G − C) / (G + C) (Wang et al. 2017). Genetic distances between each pair of the 14 core PCGs (atp6, atp8, atp9, cob, cox1, cox2, cox3, nad1, nad2, nad3, nad4, nad4L, nad5, nad6), and rps3 was calculated with MEGA 11 (Caspermeyer 2016), using the Kimura−2-parameter (K2P) model. The DnaSP v6 (Rozas et al. 2017) was used to calculate the nonsynonymous substitution rate (Ka) and the synonymous substitution rate (Ks) for all the 14 core PCGs from the 50 Hypoxylaceae specimens mitogenomes. Codon usage analysis was conducted using the Sequence Manipulation Suite (Stothard 2000), based on the genetic code 4. Genome synteny of the 50 Hypoxylaceae specimens mitogenomes and representative species from other genera was analyzed using the Mauve v2.4.0 (Darling et al. 2010).
Nuclear marker sequence alignment and phylogenetic analysis
The obtained ITS, LSU, tub2, rpb2 sequences were compared with each other and all the known sequences in GenBank using the BLAST algorithm for precise identification (https://www.ncbi.nlm.nih.gov) (Altschul et al. 1990). The molecular phylogeny was inferred from a combined dataset of ITS, tub2 and rpb2 sequences. Reference sequences were obtained from open databases and published literature, encompassing all available representative strains across the target clades, together with the closest BLASTn matches to the newly generated sequences (Supplementary Table 8).
Phylogenies were inferred under both Maximum Likelihood (ML) and Bayesian Inference (BI) frameworks for each dataset. Sequences were aligned using the MAFFT v.7.110 online program (Katoh et al. 2019) with the default settings. The alignment was manually refined using BioEdit v7.0.5.3 (Hall 1999) to correct misalignments and trim regions with excessive gaps or ambiguities. The maximum likelihood (ML) analysis was implemented in RAxML v.8.2.12 using the GTRGAMMA substitution model with 1,000 bootstrap replicates (Stamatakis 2015). Bayesian inference was performed using MrBayes v. 3.2.2 (Ronquist et al. 2012). The Markov chain Monte Carlo (MCMC) sampling in MrBayes v.3.2.2 (Ronquist et al. 2012) was used to determine the posterior probabilities (PP). Six simultaneous Markov chains were run for 1,000,000 generations, and trees were sampled every 1,000th generation. The phylogenetic tree was visualized in FIGTREE v.1.4.3 (Rambaut 2012). All analyses were run on the CIPRES Science Gateway v 3.3 web portal (Miller et al. 2010). All the obtained sequences were deposited in the GenBank (Supplementary Table 9). The alignment file is attached to this paper as supplemental material.
Mitogenome sequence alignment and mitogenome‑based phylogenetic analysis
Mitogenome-based phylogenetic analyses were performed following the methods of (Wu et al. 2021). Mitochondrial genome sequences of representative related taxa were retrieved from GenBank (https://www.ncbi.nlm.nih.gov) (Altschul et al. 1990).
The mitogenomic phylogenetic relationships of the 50 Hypoxylaceae specimens and other ascomycetes were investigated based on combined mitochondrial gene datasets (14 core PCGs). Five distinct datasets for phylogenetic analysis including concatenated nucleotide sequences of 14 conserved PCGs; PCG12 dataset comprising only the first and second codon positions of the 14 conserved genes; PCGt concatenated sequences of the 14 conserved PCGs with associated tRNAs; PCGA combining the amino acid sequences of the 14 PCGs and ITS-LSU-PCG a combined dataset of mitochondrial and nuclear markers, concatenating the sequences of the 14 conserved PCGs with two nuclear genes ITS and LSU were used (Li et al. 2020). Individual mitochondrial genes were aligned using MAFFT v7.037 software (Katoh et al. 2019), and then concatenated them into a combined mitochondrial gene dataset using SequenceMatrix v1.7.8 (Vaidya et al. 2011). A preliminary partition homogeneity test was carried out to detect potential phylogenetic conflicts between different genes. Best-fit models (GTRGAMMA substitution model ) of evolution and partitioning schemes for the gene set was determined according to Partition Finder 2.1.1 (Lanfear et al. 2017). MrBayes v3.2.6 (Ronquist et al. 2012) was used to construct the phylogenetic tree using Bayesian inference (BI) method based on the combined gene sets. Two independent runs with four chains (three heated and one cold) were conducted simultaneously for 2 × 106 generations. Each run was sampled every 1000 generations. Assumption of stationarity to reach estimated sample size (ESS) greater than 100, and the potential scale reduction factor (PSRF) approached 1.0 was checked. The first 25% samples were discarded as burn-in, and the remaining trees were used to calculate Bayesian posterior probabilities (BPP) in a 50% majority-rule consensus tree. Maximum likelihood (ML) phylogenetic trees were constructed using RAxML v8.0.0 (Stamatakis 2015). The phylogenetic tree was visualized in FIGTREE v.1.4.3 (Rambaut 2012). All analyses were run on the CIPRES Science Gateway v 3.3 web portal (Miller et al. 2010).
Comparative mitogenomic analysis and intron analysis
The genome sizes, gene numbers, intron numbers, base compositions, and gene arrangements of the 50 Hypoxylaceae specimens were analyzed to assess variations and conservation between different Hypoxylaceae species following the methods of Wu et al. (2021). The introns of cox1 genes in published Ascomycota mitogenomes were classified into different position classes (Pcls) according to the method described by Férandon et al. (2010). The cox1 genes of 50 Hypoxylaceae mitogenomes were aligned with the reference cox1 gene of the medicinal fungus Ganoderma calidophilum (Li et al. 2019), using ClustalW (Thompson et al. 1994). Introns were named according to the insertion sites (nt) in the corresponding reference gene. Each Pcl was constituted by introns inserted at the same position of the corresponding reference cox1 gene (Cheng et al. 2021). Pcls with the same insert sites were considered as orthologous introns and had high sequence similarities (Férandon et al. 2010).
Evolutionary rates of the mitochondrial genes
The synonymous substitution rate (Ks), nonsynonymous substitution rate (Ka), and their ratios (Ka/Ks) are often employed to measure evolutionary rates (Kryazhimskiy & Plotkin 2008). Therefore, we selected 14 PCGs (atp6, atp8, atp9, cob, cox1, cox2, cox3, nad1, nad2, nad3, nad4, nad4L, nad5, and nad6) and rps3 gene of 50 Hypoxylaceae specimens to calculate the values of Ka, Ks, and Ka/Ks. These genes were aligned using MEGA 11 (Tamura et al. 2021) according to the codons (parameters: Gap opening penalty: 10.00; Gap extension penalty: 0.10; and delay divergent cutoff: 30%). The Ka/Ks ratio was calculated using DnaSP v6 (Rozas et al. 2017).
Results
Phylogenetic analyses
Phylogenetic tree analysis based on combined loci of ITS, LSU, rpb2 and tub2
The final dataset included 317 sequences representing 225 taxa, comprising both newly generated sequences and those retrieved from GenBank. Following the exclusion of ambiguously aligned regions and gaps, the final data matrix comprised 2,894 characters (ITS: 1-275 bp; LSU: 276-1043 bp; rpb2: 1044-1923 bp; tub2: 1924-2894 bp). Biscogniauxia nummularia (MUCL 51395), Graphostroma platystoma (CBS 270.87), Xylaria arbuscula (CBS 126415), and X. hypoxylon (CBS 122620) were added as out-groups. The RAxML analysis of the combined ITS-LSU-rpb2-tub2 dataset yielded the best-scoring tree (Fig. 1). The tree topology (branching pattern and clade composition) from the Maximum Likelihood (ML) analysis closely resembled that of the Bayesian Inference (BI) analysis.
The phylogram is represented with 15 clades (Fig. 1). The genera Annulohypoxylon (clade 1-5), Hypomontagnella (clade 8), Jackrogersella (clade 6) and Parahypoxylon each form distinct, monophyletic clade. Hypoxylon species are distributed across multiple clades, from clade 7 to clade 15. The interposition of the genera Daldinia, Hypomontagnella, Pyrenopolyporus, Rhopalostroma, Ruwenzoria, and Thamnomyces among these clades renders the status of Hypoxylon polyphyletic. The support values for several of these clades were generally low, which further complicated the reliable resolution and clustering of Hypoxylon species.
The sequences of our collected specimens clustered within clades that align with their respective genera, supporting their morphological identifications. The phylogenetic placement of each strain is discussed in the “Notes” section for the corresponding taxon.
Phylogenetic tree based on mitochondrial genome data
The dataset comprised 50 newly generated mitochondrial genomes plus five additional sequences retrieved from GenBank, which were used as outgroups (Supplementary Tables 8–9). We constructed maximum likelihood (ML) and
bayesian Inference (BI) phylogenetic trees based on the following datasets: (1) a combined matrix of nuclear ribosomal loci (ITS, LSU) and the 14 mitochondrial protein-coding genes (PCGs); (2) the amino acid sequences of the 14 mitochondrial PCGs; (3) the nucleotide sequences of the 14 mitochondrial PCGs using only first and second codon positions; (4) the full nucleotide sequences of the 14 mitochondrial PCGs; and (5) the nucleotide sequences of the 14 mitochondrial PCGs including tRNA genes. No significant differences were observed in branching patterns or clade composition among the phylogenetic trees inferred from mitochondrial genome sequences. The phylogenetic tree based on the combined ITS–LSU–PCG dataset is presented in Fig. 2. The other phylogenetic trees are included in the supplementary documents (Supplementary Figs. 1–4).
The mitogenome phylogenetic analyses revealed strongly supported backbone nodes within Hypoxylaceae, particularly among Annulohypoxylon species, which formed monophyletic, well-resolved clades with high bootstrap support (≥90%). Hypoxylon certain nodes and clades showed lower statistical support (bootstrap values <70%), though their topological placement remained consistent across repeated analyses. The phylogenetic results supported the monophyly of Annulohypoxylon and Jackrogersella. Hypomontagnella forms a distinct monophyletic clade in phylogenies based on ITS, LSU, rpb2, and tub2 sequences (Fig. 2), but did not in the mitogenomic phylogeny, instead forming a well-supported sister branch to Hypoxylon populi. This highlights the need for broader sampling of mitogenomes to conclusively resolve the full phylogenetic classification within Hypoxylaceae. The absence of mitogenome data for many type species of key genera limits definitive conclusions, underscoring the necessity of including these taxa to resolve taxonomic relationships robustly.
Taxonomy
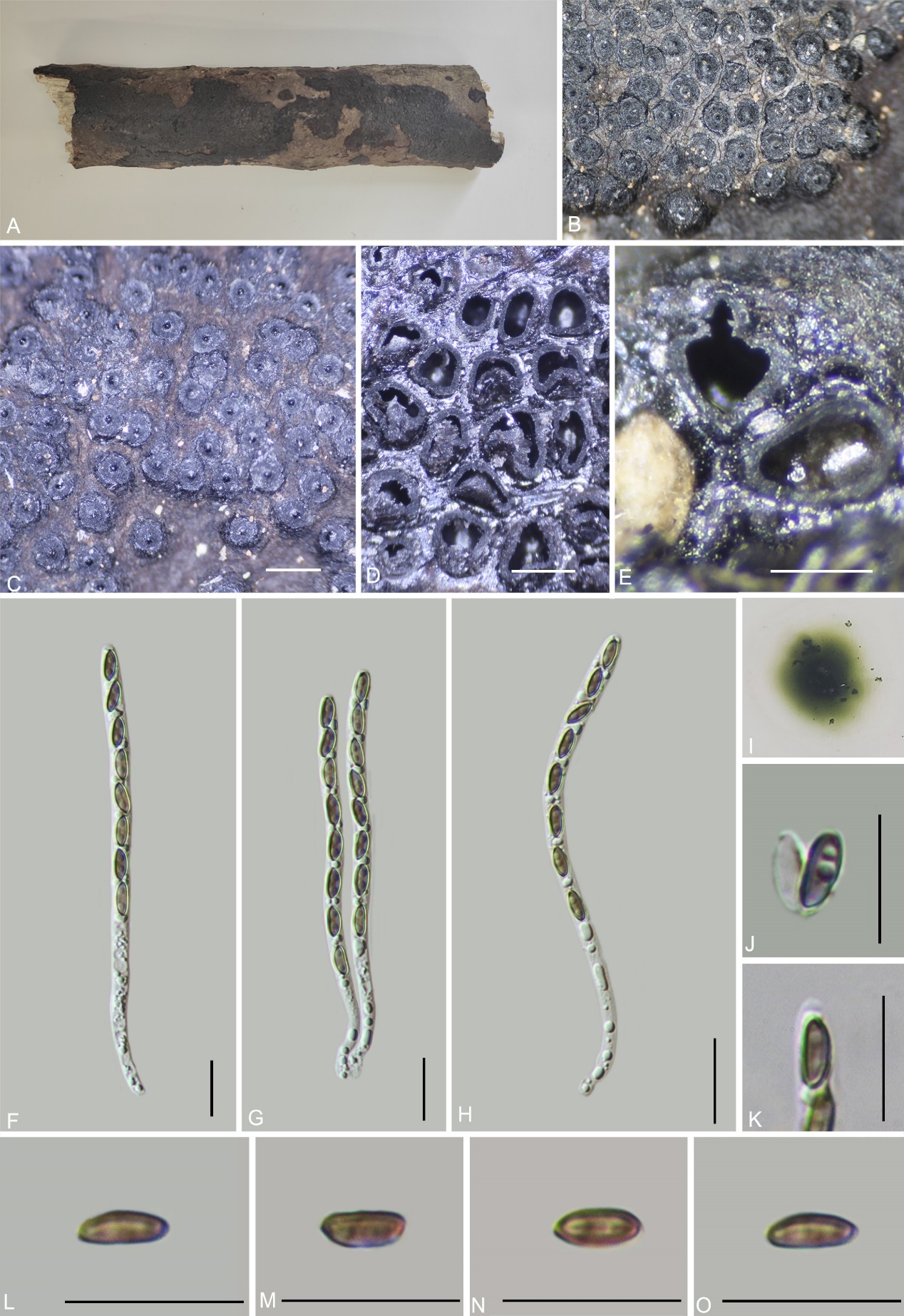
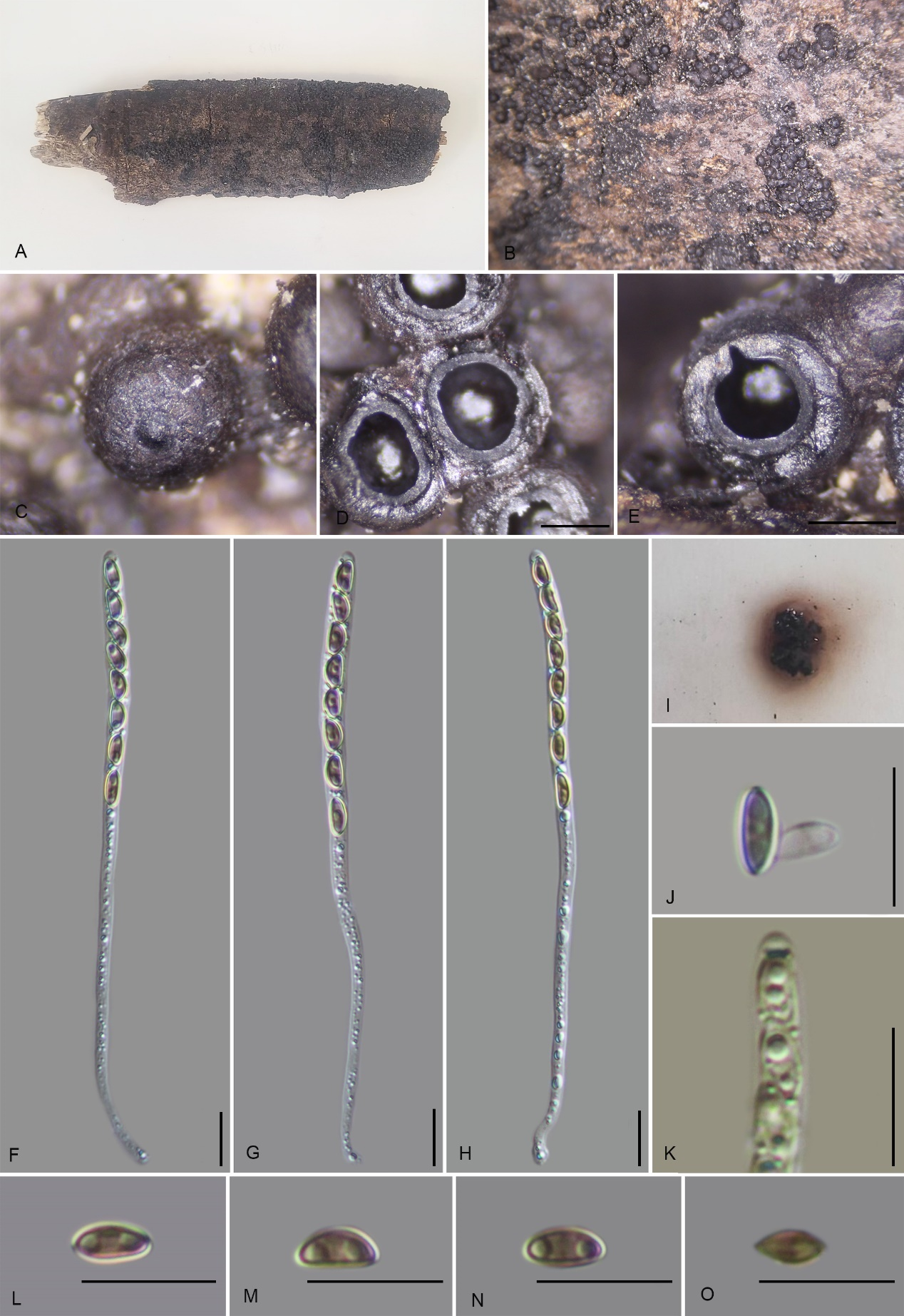
Annulohypoxylon baishaense H.M. Hu, K. Habib & Q. R. Li sp. nov.
MycoBank No.: MB858997; Fig. 3
Etymology: The epithet refers to the Baisha County, where the holotype was collected.
Material examined: China, Hainan Province, Baisha County, Yinggeling National Nature Reserve (19°33'45.80"N, 108°10'06.18"E), altitude 998 m, on dead wood, 18 July 2022, H.M. Hu, 2022YGL76 (GMB4213, holotype; GMBC4213, ex-type living culture; KUN-HKAS 148611, isotype).
Teleomorph: Stromata on the surface of dead wood, 0.4–3.5 cm long × 0.4–2 cm wide × 0.6–0.9 mm high (x̄ = 2 cm × 1.2 cm × 0.75 mm, n = 10), effused-pulvinate, hemispherical, slightly constricted at base, surface black, shiny, carbonaceous; with perithecial mounds up to 1/3 exposed, tissue below the perithecial layer black, conspicuous, with 10% KOH extractable pigments Dark Brick (66). Perithecia 1.16–1.45 mm wide × 1.39–1.66 mm high (x̄ = 1.26 × 1.49 mm, n = 20), spherical. Ostioles conical papillate, encircled with a bovei-type disc, 0.5–0.75 mm diam. Asci 160–195 × 4–6.5 µm (x̄ = 169.3 × 5.1 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, the spore-bearing part 70.8–80 µm (x̄ = 77 µm, n = 30) long, the stipes 90–115 µm (x̄ = 101 µm, n = 30) long, with apical apparatus bluing in Melzer’s iodine reagent, discoid, 2.0–2.4 × 0.9–1.5 µm (x̄ = 2.2 × 1.2 µm, n = 20). Ascospores 7.5–9 × 2.8–4.2 µm (x̄ = 8 × 3.5 µm, n = 30), uniseriate, unicellular, inequilateral ellipsoid, with rounded ends, brown, with an inconspicuous germ slit, covering the full spore length; perispore dehiscent in 10 % KOH, smooth with a thickening on the convex side, epispore smooth. Asexual morph: not formed.
Culture characteristics: Colonies grown on OA at 25 °C for 3 weeks reached a diameter of 5 cm. Colonies surface white, felty to cottony, thin; reverse light red. Not sporulating on OA nor on PDA.
Additional material examined: China, Hainan Province, Baisha County, Yinggeling National Nature Reserve (19°33'46.84"N, 108°10'04.63"E), altitude 1037 m, on dead wood. 20 July 2022, H.M. Hu, 2022YGL347 (GMB4726).
Notes: Morphologically and phylogenetically (Fig. 1), Annulohypoxylon baishaense is closely related to A. purpureonitens, sharing similarly sized ostiolar discs and ascospores. However, A. purpureonitens differs in having red-brown to black granules immediately beneath the surface (vs. black in A. baishaense), smaller ascomata (0.6–1 mm diam. vs. 1.16–1.45 mm diam.), Vinaceous Purple (101) KOH-extractable pigments [vs. Dark Brick (66)], and shorter asci (94–102.5 µm in length vs. 160–195 µm in length) (Sir et al. 2018; Cruz et al. 2021).
Phylogenetically, Annulohypoxylon baishaense is also closely related to A. kwolekiae, a species proposed based on molecular sequence data without morphological descriptions. Pairwise sequence identity between A. baishaense and A. kwolekiae (BRIP 72473a) was 92% for ITS, 100% for LSU, and 83% for rpb2. However, we do not accept A. kwolekiae as a validly published species due to the absence of a proper description, photographs, and failure to meet the standard requirements for species introduction (Tan & Shivas 2022). Further research including the acquisition of morphological data for A. kwolekiae is necessary to properly resolve its taxonomic status.
The KOH-extractable pigments of Annulohypoxylon baishaense is comparable to that of A. bahnphadengense, which yields Isabeline (65) to Grayish Sepia (106) pigmentation. However, A. bahnphadengense differs in having shorter asci (100–130 µm in total length) with a shorter stipe (37–60 µm) and is also phylogenetically distant (Fournier et al. 2010).
Other species in the clade that are morphologically similar include Annulohypoxylon nouraguense which can be distinguished by its dark vinaceous (82) to blackish brown surface, smaller ascospores (6–7 × 2.9–3.7 µm vs. 7.5–9 × 2.8–4.2 µm in A. baishaense), and Vinaceous Grey (116) to Vinaceous Purple (101) KOH-extractable pigments (Fournier & Lechat 2016).
Annulohypoxylon linjiangense H.M. Hu, K. Habib & Q.R. Li sp. nov.
MycoBank No.: MB858998; Fig. 4
Etymology: The epithet refers to the collection location, Linjiang Village.
Material examined: China, Guizhou Province, Xishui National Nature Reserve, Linjiang Village (28°13'35.9"N, 106°52'22.2"E), altitude 500 m, on dead wood of an unidentified plant, 5 Nov. 2022, H.M. Hu, 2022LJ5 (GMB4216, holotype; GMBC4216, ex-type; KUN-HKAS 148612, isotype).
Teleomorph: Stromata on the surface of dead wood, 0.1–1.5 cm long, 0.3–0.8 cm wide, and 0.5–1 cm thick, pulvinate, adnate, with perithecial mounds 1/2–3/4 exposed, carbonaceous, surface black, tissue between the perithecia black, tissue beneath perithecial layer inconspicuous, black, with 10% KOH extractable pigments Isabeline (65) to Grayish Sepia (106). Perithecia 0.84–1 mm wide × 0.76–1 mm high (x̄ = 0.87 × 0.91 mm, n = 20), spherical. Ostiole papillate, slightly lower than the level of stromatal surface,
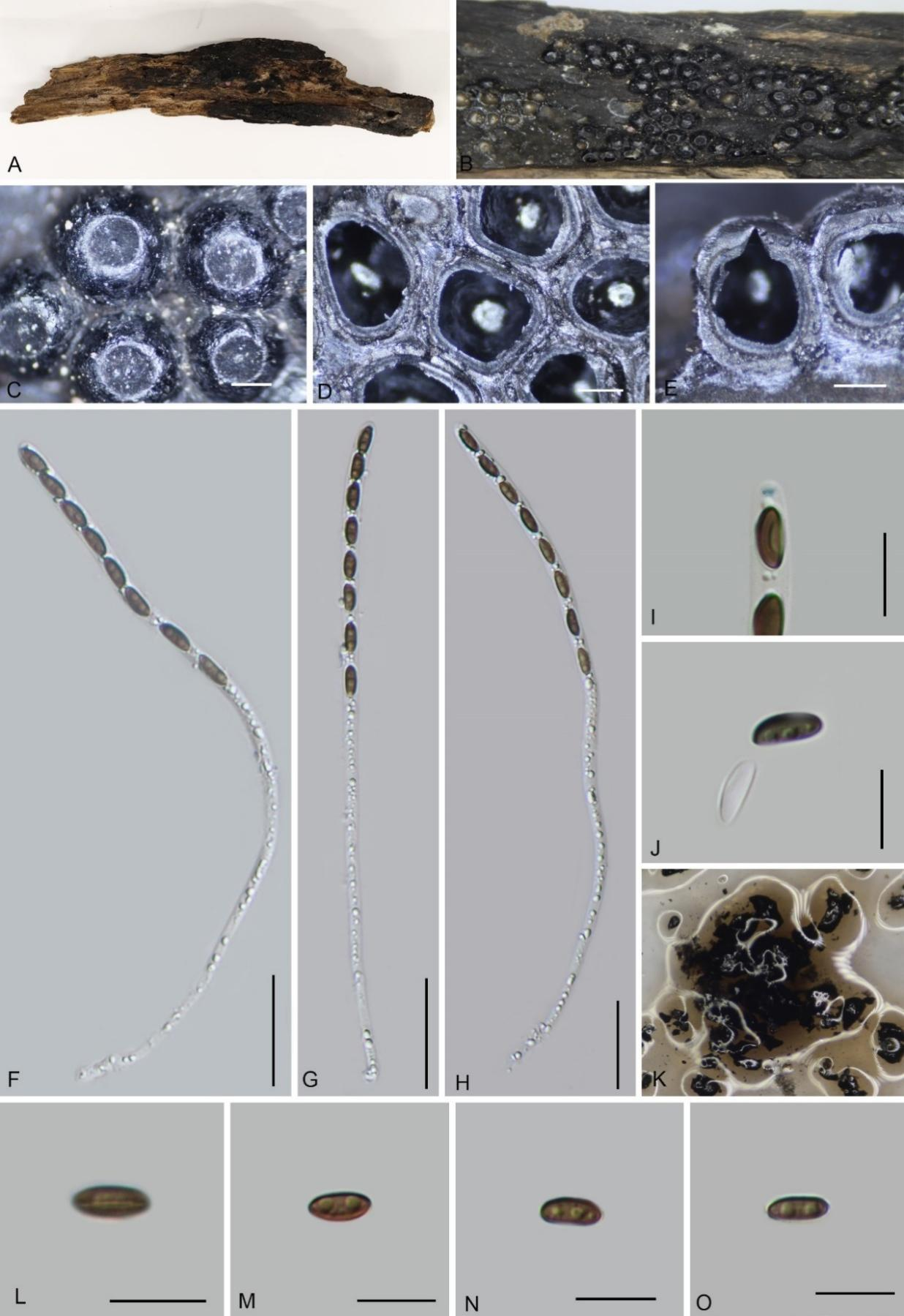

surrounded by a black disc, 0.29–0.38 mm diam. Asci 140–170 × 5.6–7 µm (x̄ = 150 × 6.6 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, the spore-bearing part 70–90 µm (x̄ = 84 µm, n = 30), apical apparatus not bluing in Melzer’s reagent. Ascospores 8–12 × 4–5.6 µm (x̄ = 10 × 4.8 µm, n = 60), uniseriate, unicellular, inequilateral ellipsoid, with rounded ends, brown, with a straight germ slit covering the full spore length, perispore dehiscent in 10 % KOH, smooth, with a thickening on the convex side, epispore smooth. Asexual morph: Not found.
Additional material examined: China, Guizhou Province, Xishui National Nature Reserve, Linjiang Village (28°13'34.1"N, 106°52'23.7"E), altitude 391 m, on dead wood of an unidentified plant, 1 Nov. 2022, H.M. Hu, 2022LJ307 (GMB4720).
Culture characteristics: Colonies grown on OA at 25 °C for 3 weeks reached a diameter of 4 cm. Colonies surface at first white become pale white, felty to cottony, thin; reverse light yellow. Not sporulating on OA nor on PDA.
Notes: Morphologically and phylogenetically, Annulohypoxylon linjiangense is closely related to A. microdiscum, sharing similar pigmentation, ascospore size, and non-amyloid apical apparatus. However, A. microdiscum can be distinguished by the presence of orange and blackish granules immediately beneath the surface and between the perithecia, much smaller asci (94–110 µm in length vs. 140–170 µm in A. linjiangense), and an indehiscent perispore (vs. dehiscent in A. linjiangense). Pairwise sequence identity between A. linjiangense and A. microdiscum was 98.87% for ITS, 94.59% for tub2 (β-tubulin), and 97.91% for rpb2.
Other phylogenetically close species, Annulohypoxylon coniformis and A. terebratum, can be easily distinguished by the absence of an ostiolar disc. Moreover, A. coniformis produces Mouse Grey (119) pigments, while A. terebratum yields Greenish Olivaceous (90) pigments in KOH (Fournier et al. 2010; Li et al. 2025).
Annulohypoxylon xishuiense H.M. Hu, K. Habib & Q.R. Li sp. nov.
MycoBank No.: MB858999; Fig. 5
Etymology: The epithet refers to the Xishui County, where the holotype was collected.
Material examined: China, Guizhou Province, Xishui County, changqiangou village (28°40'63.23"N,106°18'80.46"E), altitude 868 m, on dead branch of an unidentified plant, 4 Nov. H. M. Hu 2022CQG9 (GMB4314, holotype; GMBC4314, ex-type; KUN-HKAS 148613, isotype).
Teleomorph: Stromata on dead branch of an unidentified plant, 3–10 cm long × 1.5–5 cm wide × 0.7–1 mm high, effused to effused-pulvinate, with inconspicuous perithecial mounds, carbonaceous, surface Cinnamon (62) or Brown Vinaceous (84), gradually fading to dull Isabelline (65), black below and between perithecia, carbonaceous; tissue below the perithecial layer inconspicuous; with 10% KOH extractable pigments Greenish Olivaceous (90). Perithecia 0.36–0.83 mm wide × 0.40–0.85 mm high (x̄ = 0.55 × 0.71 mm, n = 20), spherical. Ostioles papillate, slightly higher than the level of stromatal surface, disc truncatum-type, 0.23–0.37 mm diam. Asci 85–110 × 4–5.5 µm (x̄ = 96 × 4.6 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, the spore-bearing part 47–60 µm (x̄ = 54 µm, n = 30), the stipes 40–50 µm long, apical apparatus not bluing in Melzer’s reagent. Ascospores 5.8–7 × 2.5–3.5 µm (x̄ = 6.2 × 2.8 µm, n = 30), uniseriate, unicellular, ellipsoid, almost equilateral, with rounded ends, brown, with a straight spore-length germ slit, perispore dehiscent in 10 % KOH, smooth, with a thickening on the convex side, epispore smooth. Asexual morph: not formed.
Culture characteristics: Colonies grown on OA at 25 °C for 4 weeks reached a diameter of 5 cm. Colonies surface at first white, felty to cottony, thin; reverse greyish white. Not sporulating on OA nor on PDA.
Additional material examined: China, Guizhou Province, Xishui County, changqiangou village (28°40'61.58"N, 106°18'79.28"E), altitude 902 m, on dead branch of an unidentified plant, 4 Nov. H.M. Hu 2022CQG313 (GMB4725).
Notes: Morphologically, Annulohypoxylon xishuiense closely resembles A. maolanense in stromatal surface coloration, KOH-extractable pigmentation, and the size of asci and ascospores. Phylogenetically, the two species also appear as sister taxa, albeit with low statistical support. A key phenotypic difference is the larger ostiolar disc in A. xishuiense (0.23–0.37 mm) compared to A. maolanense (0.15–0.2 mm). Moreover, A. maolanense has smaller perithecia (0.25–0.36 mm wide) than those of A. xishuiense (0.36–0.83 mm wide) (Li et al. 2025).
The stromata of A. xishuiense could be confused with A. stygium. However, A. xishuiense can be easily distinguished by its much larger ostiolar discs (0.23–0.37 mm vs. 0.1–0.2 mm in A. stygium). Additionally, Ju & Rogers (1996) and Fournier & Lechat (2016) reported the ascospores of A. stygium as ellipsoid-inequilateral, whereas the ascospores of the new species are almost equilateral. Based on these morphological differences and phylogeny, we propose A. xishuiense as a new species.
Hypomontagnella vinacea K. Habib, Y.H. Pi & Q.R. Li sp. nov.
MycoBank No.: MB859000; Fig. 6
Etymology: The epithet refers to the vinaceous color of the stromatal surface.
Material examined: China, Guizhou Province, Rongjiang County, Leigongshan Nature Reserve (26°22'54.31"N, 108°21'51.63"E), altitude 923 m, on dead branches of an unidentified plant, 29 Aug. 2020, Yinhui Pi, 2020LGS48 (GMB0247, holotype; GMBC0247, ex-type; KUN-HKAS 148614, isotype).
Teleomorph: Stromata on the surface of dead wood, 1.7–2.8 cm long × 0.8–1.5 cm wide × 0.5–0.65 mm high (x̄ = 2.0 cm × 0.94 cm × 0.62 mm, n = 10), effused to effused-pulvinate, with inconspicuous perithecial mounds, surface dark Vinaceous Grey (116) or blackish, carbonaceous, tissue between perithecia black, tissue below the perithecial layer inconspicuous, without pigments in 10 % KOH. Perithecia 0.34–0.58 mm wide × 0.51–0.81 mm high (x̄ = 0.44 × 0.69 mm, n = 20), spherical to obovate. Ostioles papillate, higher than the level of the stromatal surface, without a disc. Asci 80–145 × 4.5–7 µm (x̄ = 111 × 6 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, spore-bearing part 45–55 µm (x̄ = 50 µm, n = 30) long, stipes 55–75 µm (x̄ = 57 µm, n = 30) long, apical apparatus amyloid, discoid, 1.3–2.3 µm wide × 0.9–1.8 µm high (x̄ = 1.8 × 1.2 µm, n = 20). Ascospores 7.5–8.5 × 3–4.5 µm (x̄ = 7.8 × 3.6 µm, n = 30), uniseriate, unicellular, ellipsoidal, inequilateral, with narrowly rounded ends, brown,
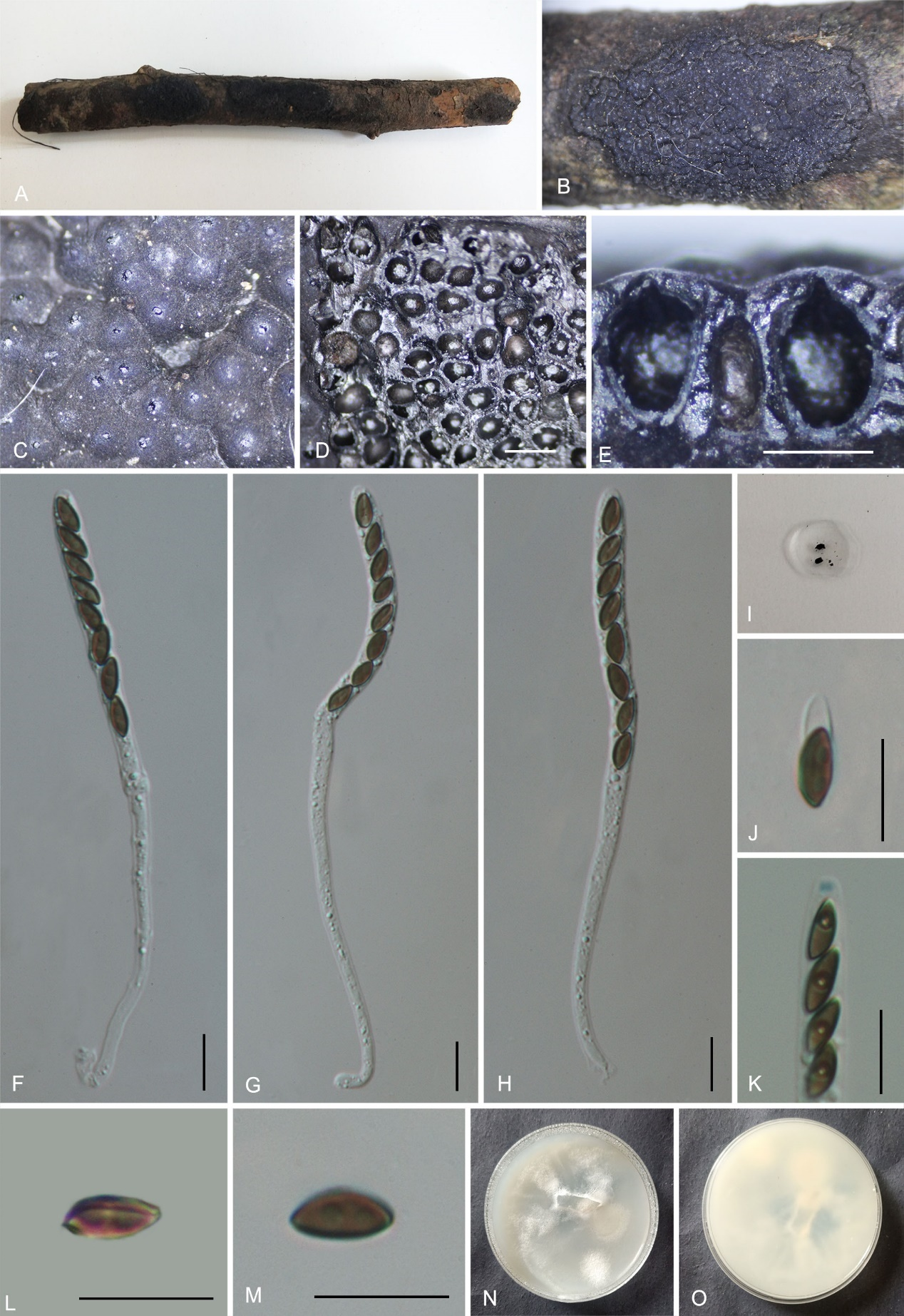
with an inconspicuous straight spore-length germ slit; perispore always dehiscent in 10% KOH, smooth, epispore smooth. Asexual morph: not formed.
Culture characteristics: Colonies on OA covering 4–5 cm diam. after 2 weeks at 25 °C, Colonies surface white, cottony, azonate with entire margins; after 4 weeks covering a 9 cm petri dish, becoming white grey, appressed with entire margins; reverse white. No conidia were observed.
Additional material examined: China, Guizhou Province, Libo County, Qiannan Buyi and Miao Autonomous Prefecture, Maolan National Nature Reserve (25°18'2.76"N, 108°4'29.48"E), altitude 545 m, on dead branches of an unidentified plant, 9 July 2021, Y.H. Pi, 2021MLB37 (GMB0248), living culture GMBC0248.
Notes: Morphologically, Hypomontagnella vinacea resembles H. monticulosa, sharing similar stromatal coloration. However, H. monticulosa is distinguished by a conspicuous disc surrounding the ostioles, larger ascospores (x̄ = 10.4 × 4.8 μm) with a sigmoid germ slit, and young stromata produce Vinaceous Purple (101) pigments in KOH (Ju & Rogers 1996; Lambert et al. 2019). Pairwise sequence identity between H. vinacea (GMB0247) and H. monticulosa (MUCL 54604) was 96.4% for ITS, 99.6% for LSU, 93.7% for rpb2, and 96.9% for tub2. Hypomontagnella barbarensis also shares a vinaceous stromatal surface but differs in having much larger ascospores (13–19.3 × 6.9–9.4 µm vs. 7.5–8.5 × 3–4.5 µm) and ostiole surrounded by a disc. Hypomontagnella vinacea is distinguished from all known species in the genus by its smaller ascospores (7.5–8.5 × 3–4.5 µm) and the absence of a disc surrounding ostiole, whereas other species in the genus typically have ascospores longer than 9 µm and broader than 4.5 µm (Lambert et al. 2019).
Hypomontagnella yinggelingensis H.M. Hu, K. Habib & Q.R. Li sp. nov.
MycoBank No.: MB859001; Fig. 7
Etymology: The epithet refers to Yinggeling National Nature Reserve, where the holotype was collected.
Material examined: China, Hainan Province, Baisha County, Yinggeling National Nature Reserve (18°28'59.1"N, 109°20'26.2"E), altitude 1088 m, on dead twig of an unknown plant, 18 July 2022, H.M. Hu, 2022YGL39 (GMB4715, holotype; GMBC4715, ex-type; KUN-HKAS 148615, isotype)
Teleomorph: Stromata on the surface of dead wood, 0.3–6 cm long × 0.3–0.8 cm wide × 0.85–1 mm thick, cushion-shaped, surface black, carbonaceous, the tissue between perithecia black, tissue below the perithecial layer inconspicuous, with 10% KOH extractable pigments Vinaceous Purple (101). Perithecia 0.37–0.60 mm wide × 0.62–0.77 mm high (x̄ = 0.49 × 0.68 mm, n = 20), obovate. Ostioles conical papillate, slightly higher than stromatal surface, without a disc. Asci 120–140 × 5–7 µm (x̄ = 131 × 6 µm, n = 30), 8-spored, unitunicate, cylindrical, spore-bearing part 50–65 µm long (x̄ = 56 µm, n = 30), apical apparatus bluing in Melzer’s reagent, discoid, 1.5–1.8 × 0.6–1 µm (x̄ = 1.6 × 0.8 µm, n = 20). Ascospores 7–8.4 × 3–4.3 µm (x̄ = 7.8 × 3.8 µm, n = 30), uniseriate, unicellular, ellipsoid, inequilateral, brown, with narrowly rounded ends, with a sigmoid germ slit of spore-length; perispore dehiscent in 10 % KOH, smooth, epispore smooth. Asexual morph: Not observed.
Culture characteristics: Colonies on OA covering 5 cm diam. after 3 weeks at 25 °C, Colonies surface white, cottony, azonate, margin thin; after 5 weeks covering a 9 cm petri dish, becoming yellowish, dense; reverse brownish grey. No conidia were observed.
Additional material examined: China, Hainan Province, Baisha County, Yinggeling National Nature Reserve (18°28'58.9"N, 109°20'26.3"E), altitude 1315 m, on dead twig of an unknown plant, 18 July 2022, H.M. Hu, 2022YGL327 (GMB4724).
Notes: Phylogenetically and morphologically, Hypomontagnella yinggelingensis is closely related to the above described Hy. vinacea. However, Hy. vinacea differs from Hy. yinggelingensis by its Vinaceous Grey (116) to blackish surface coloration (vs. black), without KOH extractable pigment (vs. Vinaceous Purple) and ascospores with a straight germ slit (vs. sigmoid germ slit) (this study).
Hypomontagnella yinggelingensis shares the presence of a sigmoid germ slit with Hy. monticulosa and Hy. submonticulosa. However, both species can be readily distinguished by their larger ascospores (H. monticulosa 7–11 × 3.5–4.5 µm; Hy. submonticulosa 9–13.5 × 4–5 μm). Hypomontagnella monticulosa further differs by having a vinaceous surface in young stromata, becoming brown-vinaceous to blackish in mature stromata, whereas Hy. yinggelingensis has a black stromatal surface in both immature and mature stages. Hypomontagnella submonticulosa further differs by having a straight to slightly oblique germ slit shorter than the spore length, while H. yinggelingensis has a spore-length sigmoid germ slit (Lambert et al. 2019).
Hypoxylon betulae K. Habib, Y.H. Pi & Q.R. Li sp. nov.
MycoBank No.: MB859002; Fig. 8
Etymology: The epithet refers to the host genus Betula sp.
Material examined: China, Guizhou Province, Kuankuoshui National Nature Reserve (28°31'54.18"N, 107°10'36.72"E), altitude 1556 m, on dead branches of Betula sp., 2 Nov. 2022, H.M. Hu, 2022KKS123 (GMB4231, holotype; GMBC4231, ex-type living culture; KUN-HKAS 148616, isotype).
Teleomorph: Stromata on dead branches of Betula sp., 0.9–3.5 cm long × 0.5–1.4 cm wide × 0.7–1 mm high, flat, surface Vinaceous Purple (101) or purplish grey (128), tissue black between perithecia; tissue below the perithecia dark brown to black, with 10% KOH extractable pigments Sienna (8) to Dark Brick (86). Perithecia 0.22–0.42 mm wide × 0.35–0.55 mm high (x̄ = 0.33 × 0.46 mm, n = 20), spherical. Ostiole recessed below the surface, not prominent. Asci 110–140 × 6.5–8.5 µm (x̄ = 127 × 7.3 µm, n = 30), 8-spored, unitunicate, long-cylindrical, long-stipitate, the spore-bearing part 40–50 µm long, the stipes 70–90 µm long, with apical apparatus bluing in Melzer’s iodine reagent, discoid, 1.7–3 × 1–1.5 µm (x̄ = 2.4 × 1.2 µm, n = 20). Ascospores 6–8.2 × 2.6–4.7 µm (x̄ = 7.2 × 3.6 µm, n = 30), uniseriate, unicellular, ellipsoid-inequilateral, with rounded ends, light brown to brown, with an inconspicuous germ slit; perispore dehiscent in 10% KOH. epispore smooth. Asexual morph: not formed.
Culture characteristics: Colonies growing on OA at 25 °C for 3 weeks reached a diam. of 5 cm. Colony surface white, felty, flocculent, circular, dense; reverse white, center cyan. Not sporulating on OA nor on PDA.
Additional material examined: China, Guizhou Province, Kuankuoshui National Nature Reserve (28°31'53.58"N, 107°10'34.69"E), altitude 1494 m, on dead branches of Betula sp., 5 Nov. 2022, H.M. Hu, 2022KKS307 (GMB0445).
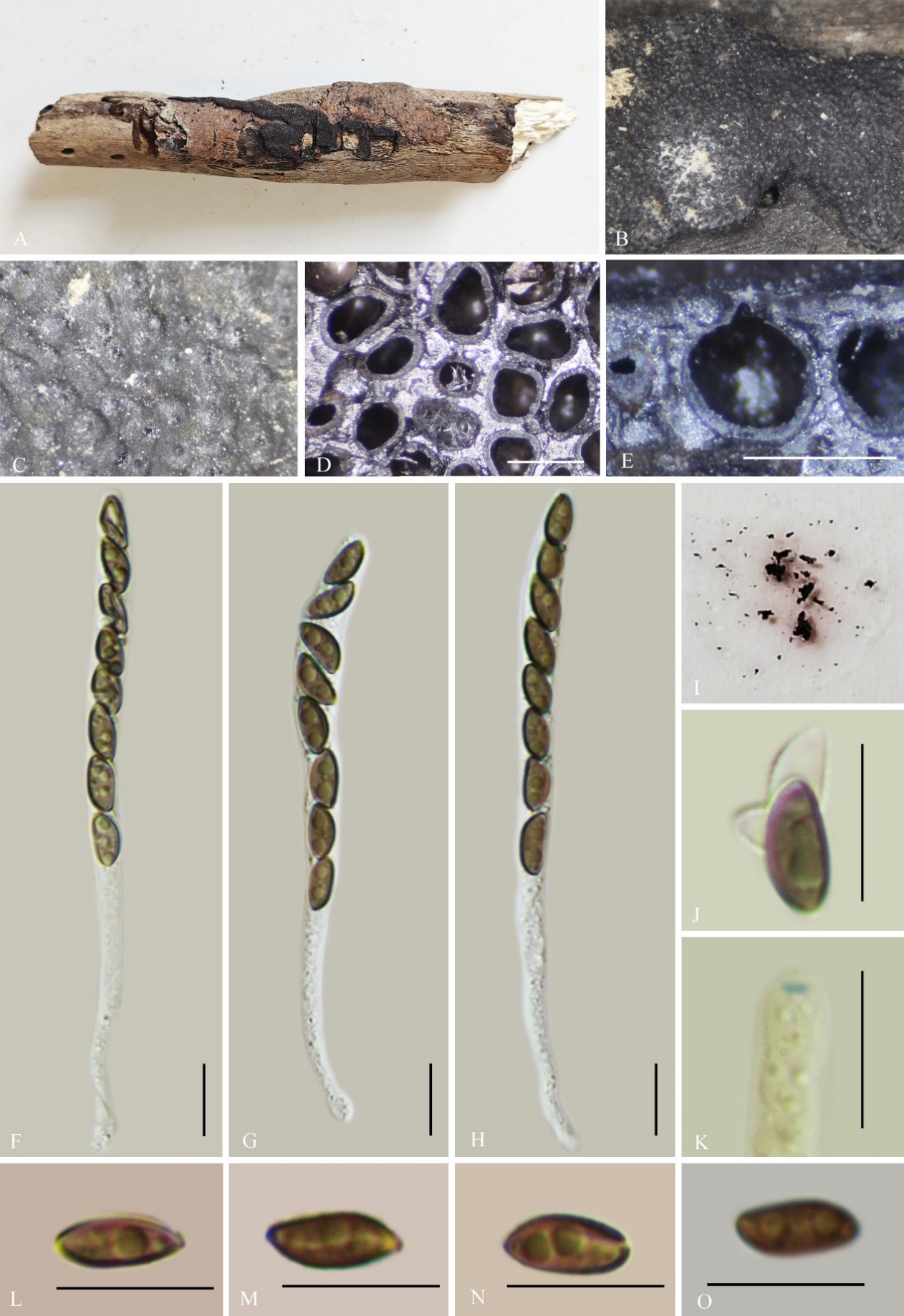
Notes: Phylogenetically (Fig. 1), Hypoxylon betulae is closely related to H. fuscopurpureum, and morphologically also resembles it, with both species sharing a purplish stromatal surface. However, H. fuscopurpureum can be differentiated by olivaceous KOH-extractable pigments, larger perithecia (0.3–0.6 × 0.22–0.41 mm), and larger ascospores (12–15 × 5.5–7 µm vs. 6–8.2 × 2.6–4.7 µm), and have an indehiscent perispore (Ju & Rogers 1996; Mühlbauer et al. 2002).
Its stromata could be confused with H. fuscum, as both species have a purplish stromatal surface. However, H. fuscum is distinguished by Amber (47), Isabelline (65), Olivaceous (48), Gray Olivaceous (107), or Greenish Olivaceous (90) KOH-extractable pigments, and larger ascospores (8–20 × 4–8 μm) (Ju & Rogers 1996; Mühlbauer et al. 2002).
Hypoxylon betulae can be distinguished from other Hypoxylon species that produce Sienna (8) stromatal KOH-extractable pigments, such as H. erythrostroma (asci 90–115 μm in total length, ascospores with acute ends and a sigmoid germ slit), H. fendleri (asci 86–156 μm in total length, ascospores 9–12 × 4–5.5 μm, with a sigmoid germ slit), H. lateripigmentum (asci 85–155 μm in total length, with olivaceous-yellow granules beneath the surface and between the perithecia), H. musceum (asci 83–110 μm in total length, ascospores 9–13 × 4.5–6 μm), H. sepiaceum (subperithecial tissue olivaceous-brown), and H. arawakianum (asci 100–110 μm in total length, ascospores 9.7–11.2 μm) (Rogers 1981; Kuhnert et al. 2014; Fournier & Lechat 2015).
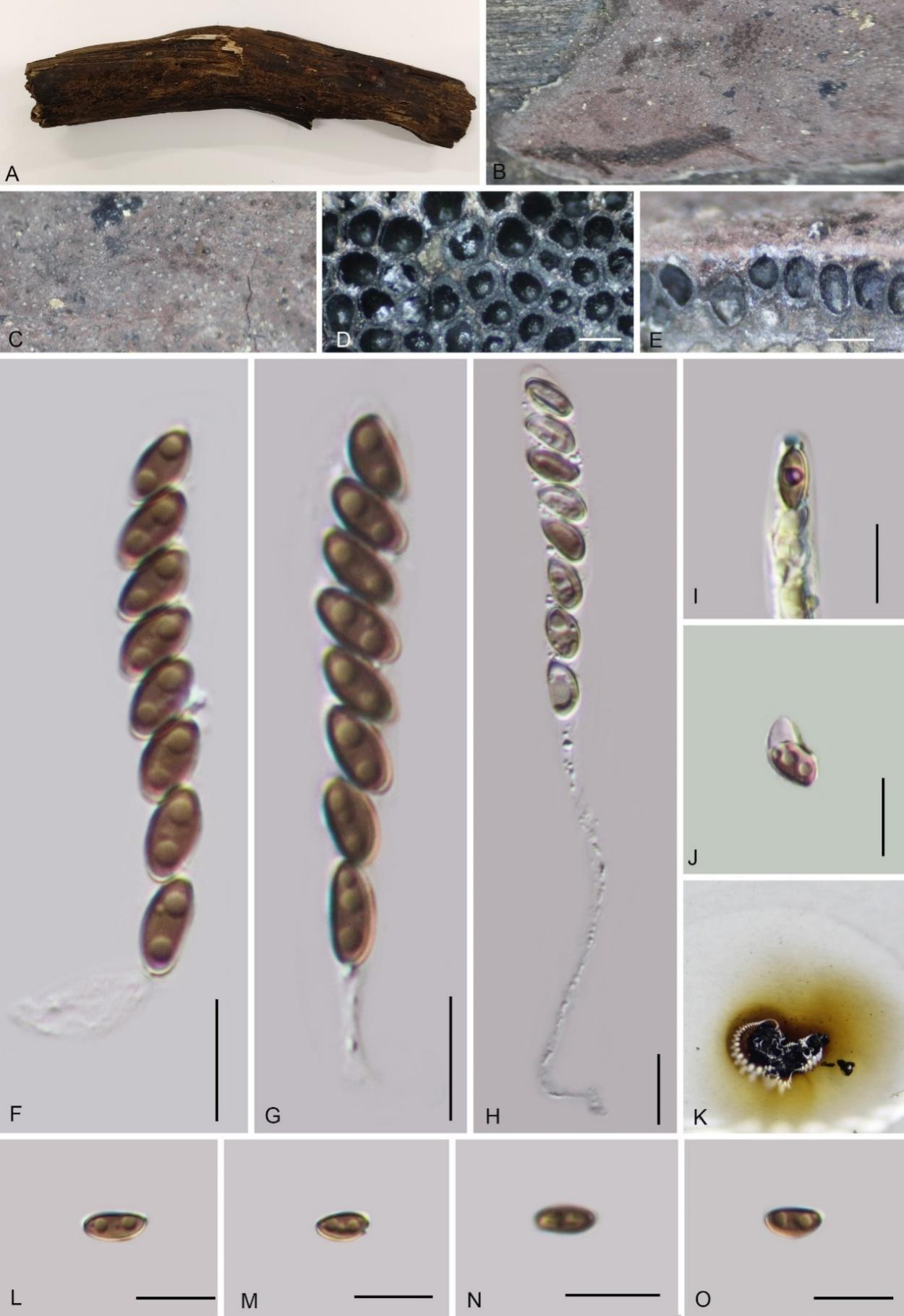
Hypoxylon guiense W.Y. Zeng, K. Habib & Q.R. Li sp. nov.
MycoBank No.: MB859003; Fig. 9
Etymology: The epithet refers to "Gui" a local name for Guangxi Zhuang Autonomous Region, where the holotype was collected.
Material examined: China, Guangxi Zhuang Autonomous Region, Qinzhou City, Shiwandashan National Nature Reserve, (21°25'45.5"N, 107°11'29.8"E), altitude 650 m, on dead culm of bamboo, 14 Aug. 2023, W.Y. Zeng, 2023SWDS30 (GMB4712, holotype; ex-type GMBC4712; KUN-HKAS 148617, isotype)
Teleomorph: Stromata on the surface of dead culm of bamboo, 0.4–1.2 cm long × 0.2–0.8 cm wide × 0.2–1.2 mm thick, effused-pulvinate, surface Dark Brick (66) to Brown Vinaceous (84), tissue between the perithecia black, tissue below the perithecial layer blackish brown, with KOH-extractable pigments Dark Olivaceous Grey (121), Umber (9) to Sepia (63) after a while. Perithecia 0.37–0.46 mm wide × 0.38–0.44 mm high (x̄ = 0.42 × 0.42 mm, n = 20), subglobose. Ostiole lower than the surface, inconspicuous. Asci 121–160 × 9–11 µm (x̄ = 141 × 10.1 µm, n = 30), 8-spored, long cylindrical, spore-bearing part 97–126 µm long (x̄ = 106 µm, n = 30), with apical apparatus bluing in Melzer’s reagent, discoid, 3.3–3.8 × 0.7–1.1 µm (x̄ = 3.5 × 0.8 µm, n = 20). Ascospores 15–20 × 7–10 µm (x̄ = 17.2 × 8.1 µm, n = 30), brown to dark brown, ellipsoid, equilateral, with rounded ends, with inconspicuous straight germ slit, 1/3–1/2 of spore-length, perispore indehiscent in 10% KOH; epispore smooth. Asexual morph: not formed.
Culture characteristics: Colonies grown on OA at 25 °C for 3 weeks reached a diameter of 5 cm. Colonies surface white, felty, cottony, thin; reverse white. Not sporulating on OA nor on PDA.
Additional material examined: China, Guangxi Zhuang Autonomous Region, Qinzhou city, Shiwandashan National Nature Reserve (21°25'44.4"N, 107°11'29.4"E), altitude 739 m, on dead culm of bamboo, 16 Aug. 2023, W.Y. Zeng, 2023SWDS308 (GMB4713).
Notes: Morphologically, Hypoxylon guiense closely resembles H. wuzhishanense, which has also been reported on bamboo and has similarly sized ascomata. However, H. wuzhishanense can be distinguished by its Pale Vinaceous (85) to Livid Vinaceous (83) and Vinaceous Purple (101) KOH-extractable pigments, shorter asci (70–139 µm total length vs. 121–160 µm), and smaller dehiscent ascospores (6.1–9.6 × 3.2–5 µm vs. indehiscent 15–20 × 7–10 µm) (Ma et al. 2022).
Morphologically, H. guiense also resembles H. pilgerianum, which has been reported on bamboo and shares similar surface coloration. Phylogenetically, the two species are also closely related. However, H. pilgerianum can be differentiated by the presence of dull orange-brown or dull reddish-brown granules immediately beneath the surface and between the perithecia, Ochreous (44), Honey (64), or Amber (47), KOH-extractable pigments and much shorter asci (68–97 µm total length) and ascospores (8–9.7 × 4–4.8 µm) (Ju & Rogers 1996).
Hypoxylon huangjingense H.M. Hu, K. Habib & Q.R. Li sp. nov.
MycoBank No.: MB859005; Fig. 10
Etymology: The epithet refers to Huangjing Old Forest Nature Reserve, where the holotype was collected.
Material examined: China, Sichuan Province, Huangjing Old Forest Nature Reserve, (28°8'6.9"N, 105°26'38.9" E), altitude 1100 m, on dead wood, 16 Aug. 2022, H.M. Hu, 2022HJLL34 (GMB4228, holotype; GMBC4228, ex-type; KUN-HKAS 148618, isotype)
Teleomorph: Stromata on the surface of dead wood, 1.5–6 cm long × 0.5–1 cm wide × 0.25–0.28 mm high, effused to effused-pulvinate, with inconspicuous perithecial mounds, slightly carbonaceous or cortical, surface Mouse Grey (118), tissue between the perithecia black, tissue below the perithecial layer conspicuous, black; with 10% KOH extractable pigments Grey Olivaceous (107) to Olivaceous Grey (121). Perithecia 0.42–0.63 mm wide × 0.55–0.81 mm high (x̄ = 0.52 × 0.70 mm, n = 20), spherical. Ostioles umbilicate, slightly higher than the surface, inconspicuous. Asci 120–140 × 5–7.5 µm (x̄ = 131.8 × 6.2 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, the spore-bearing part 60–68 µm (x̄ = 63.7 µm, n = 30), with apical apparatus bluing in Melzer’s reagent, discoid, 2–2.3 × 1.3–1.6 µm (x̄ = 2.2 × 1.4 µm, n = 20). Ascospores 6.5–9 × 3.5–5 µm (x̄ = 7.8 × 4.3 µm, n = 30), uniseriate, unicellular, brown, ellipsoidal, equilateral, with a faint straight germ slit, slightly less than spore-length, perispore dehiscent in 10 % KOH, smooth; epispore smooth. Asexual morph: Not observed.
Culture characteristics: Colonies on OA covering 3 cm diam. after 2 weeks at 25 °C, Colonies surface white, cottony, azonate, margin entire; after 6 weeks covering a 9 cm petri dish, white, dense; reverse yellow. No conidia were observed.
Additional material examined: China, Sichuan Province, Huangjing Old Forest Nature Reserve, (28°8'6.5"N, 105°26'38.3"E), altitude 1271 m, on dead wood, 19 Aug. 2022, H.M. Hu, 2022HJLL309 (GMB4721).

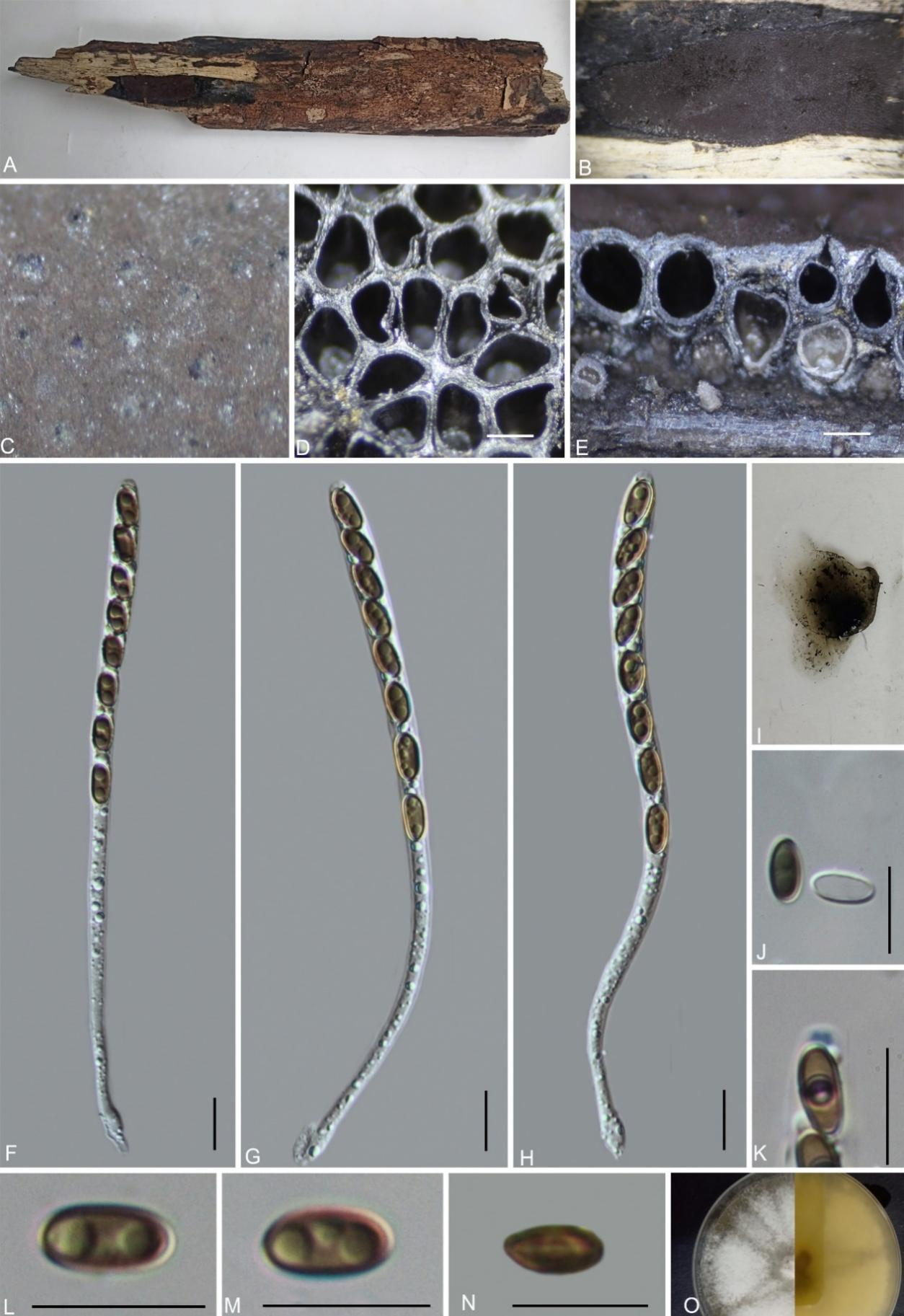
Notes: Morphologically and phylogenetically (Fig. 1), Hypoxylon huangjingense is closely related to H. investiens, sharing similarly sized asci and ascospores with a germ slit slightly less than the spore length. However, H. investiens can be distinguished by its tubular to long-tubular ascomata (0.3–0.4 mm diam × 0.5–1 mm high vs. spherical 0.42–0.63 mm wide × 0.55–0.81 mm high), Greenish Yellow (16), Dull Green (70), or Dark Green (21) KOH-extractable pigments (vs. Grey Olivaceous [107] to Olivaceous Grey [121]), and its indehiscent perispore (vs. dehiscent) (Ju & Rogers 1996).
Morphologically, H. huangjingense also resembles H. lenormandii (= H. riograndense) in having the same surface coloration and spherical perithecia. However, H. lenormandii differs by its Hazel (88), Sienna (8), Cinnamon (62), Fulvous (43), Umber (9), or Ochreous (44) KOH-extractable pigments, and larger asci (123–170 µm total length vs. 120–140 µm total length) and ascospores (9.5–16 µm length vs. 6.5–9 µm length) (Ju & Rogers 1996).
Hypoxylon lancangense K. Habib, Y.H. Pi & Q.R. Li sp. nov.
MycoBank No.: MB859006; Fig. 11
Etymology: The epithet refers to the location of the type specimen, Lancang River.
Material examined: China, Yunnan Province, Baoshan City, Lancangjiang Nature Reserve (24°57'26.28"N, 99°44'23.08"E), altitude 1314 m, on dead branches of an unidentified plant, 2 Oct. 2019, Y. H. Pi, 2019LC110 (GMB0284 holotype; ex-type living culture GMBC0284; KUN-HKAS 148619, isotype).
Teleomorph: Stromata on the surface of dead wood, 1–5 cm long × 2–4 cm wide × 0.4–1 mm thick (mean = 3.6 cm × 3.2 cm × 0.76 mm, n = 10), effused or effused-pulvinate, not carbonaceous, surface Brown Vinaceous (84), Umber (9), orange granules beneath the surface and between perithecia, with 10% KOH extractable pigments Ochreous (44) or Luteous (12) with Citrine (13) tone. Perithecia 0.24–0.58 mm wide × 0.32–0.61 mm high (x̄ = 0.43 × 0.52 mm, n = 20), tubular. Ostioles umbilicate, lower than the stromatal surface, overlain with conspicuous white substance. Asci 95–130 × 8–9 µm (x̄ = 114 × 8.5 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, spore-bearing part 65–90 µm (x̄ = 76 µm, n = 30) long, stipes 30–75 µm (mean = 50 µm, n = 30) long, with apical apparatus bluing in Melzer’s reagent, discoid, 2–2.5 µm broad × 0.7–1.4 µm high (x̄ = 2.1 × 0.9 µm, n = 20). Ascospores 9–10.5 × 4–5 µm (x̄ = 10 × 4.7 µm, n = 30), uniseriate, unicellular, inequilateral ellipsoidal with rounded ends, brown to black brown, with a faint straight germ slit nearly spore-length; perispore dehiscent in 10 % KOH, epispore smooth.
Anamorph on OA: Conidiophores Virgariella-like, hyaline, smooth. Conidiogenous cells hyaline, smooth, 63–81 × 2.3–4.5 µm. Conidia hyaline, smooth to finely roughened, ovoid, 3.5–5.5 × 2.4–3.2 µm.
Culture characteristics: Colonies grown on OA at 25 °C for 3 weeks reached a diameter of 5 cm. Colonies surface white, azonate, velvety to felty, irregular entire margin; reverse pinkish white to hazel.
Additional materials examined: China, Guizhou Province, Guiyang City, Pingba (26°25'10.24"N, 106°24'25.21"E) altitude: 1052 m, on dead branches of an unidentified plant, 5 Sept. 2020, Y. H. Pi, 2020PB23 (GMB0286, culture GMBC0286). China, Hunan Province, Yiyang City, Liubuoxi National Nature Reserve (28°23'53.73"N, 110°51'36.25"E), altitude 836 m, on dead branches of an unidentified plant, 26 Aug. 2023, W. Y. Zeng, 2023LBX41 (GMB4249; culture GMBC4249).
Notes: Morphologically, Hypoxylon lancangense is similar to H. perforatum in having the same surface coloration, ascomata overlain with a conspicuous white substance, and similarly sized asci. However, H. perforatum differs by its spherical ascomata (0.1–0.3 mm diam vs. tubular, 0.24–0.58 mm wide), smaller ascospores (8–13 × 4–6 µm vs. 9–10.5 × 4–5 µm), and larger conidia (3.5–4.8 × 2.8–3.5 µm vs. 3.5–5.5 × 2.4–3.2 µm) (Ju & Rogers 1996). Additionally, in the epitype description of H. perforatum, the ascospores are reported as larger (10–12.2 × 3.6–4.7 µm), while the asci are smaller (77.8–95.4 × 6.4–7.8 µm) compared to those of H. lancangense.
In surface coloration, ascospore size, and KOH-extractable pigments, H. lancangense is similar to H. rubiginosum. However, H. rubiginosum differs in having spherical to obovoid, larger ascomata (300–650 µm diam × 450–800 µm high vs. 0.24–0.58 mm wide × 0.32–0.61 mm high), larger asci (130–180 µm total length vs. 95–130 µm total length), and a Nodulisporium-like anamorph with larger conidia (5–6 × 3–4 µm vs. Virgariella-like anamorph conidia, 2.4–3.2 × 3.5–5.5 µm) (Ju & Rogers 1996).
In surface coloration, Virgariella-like anamorph, and ascospore size, H. lancangense is similar to H. petriniae. However, H. petriniae differs by its spherical to obovoid perithecia (vs. tubular), slightly longer asci (115–145 µm total length vs. 95–130 µm total length), and larger conidia (7–8 × 3–4 µm vs. 3.5–5.5 × 2.4–3.2 µm) (Stadler et al. 2004). In the phylogram, H. lancangense is also phylogenetically distant from these taxa.
Hypoxylon populi K. Habib, Y.H. Pi & Q.R. Li sp. nov.
MycoBank No.: MB859007; Fig. 12
Etymology: The epithet refers to the host plant genus “Populus”.
Material examined: China, Guizhou Province, Qiannan Buyi and Miao Autonomous Prefecture, Luodian County, Hongshui River (25°65'28.95"N, 107°30'21.74"E), altitude 480 m), on dead branches of Populus sp. 12 Sept. 2021, Y.H. Pi, 2021HSH7 (GMB4232, holotype; GMBC4232, ex-type; KUN-HKAS 148620, isotype).
Teleomorph: Stromata on the surface of dead branches of Populus sp., 0.6–1 cm long × 0.2–1 cm wide × 0.7–0.9 mm thick (x̄ = 0.8 cm × 0.4 cm × 0.78 mm, n = 10), glomerate, coalescent into larger pulvinate stromata, with conspicuous to inconspicuous perithecial mounds, surface dark vinaceous (82) to brown vinaceous (84); tissue between the perithecia black, tissue below the perithecial layer conspicuous, blackish brown, with 10% KOH extractable pigments Umber (9), Dark Purple (36). Perithecia 0.37–0.46 mm wide × 0.40–0.64 mm high (x̄ = 0.41 × 0.51 mm, n = 20), spherical. Ostioles inconspicuous. Asci 104–125 × 8–9.5 µm (x̄ = 113 × 9.2 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, spore-bearing part 60–90 µm (x̄ = 81 µm, n = 30) long, with apical apparatus bluing in Melzer’s reagent, discoid, 1.6–2.1 × 0.5–0.8 µm (x̄ = 2 × 0.6 µm, n = 20). Ascospores 9.5–13 × 4.5–6.6 µm (x̄ = 11.7 × 5.9 µm, n = 30), uniseriate, unicellular, dark brown, inequilateral, with rounded ends, with inconspicuous straight spore-length germ slit; perispore dehiscent in 10% KOH, with conspicuous coil-like ornamentation; epispore smooth. Asexual morph: not formed.


Additional material examined: China, Guizhou Province, Qiannan Buyi and Miao Autonomous Prefecture, Luodian County, Hongshui River (25°65'29.42"N, 107°30'23.38"E), altitude 514 m, on dead branches of Populus sp. L., 14 Sept. 2021, Y.H. Pi, 2021HSH300 (GMB0456).
Notes: Phylogenetically, Hypoxylon populi is closely related to H. lienhwacheense, which can be easily differentiated by Dark Violet (33), Livid Violet (79), or Dark Slate Blue (11) KOH-extractable pigments [vs. Umber (9) to Dark Purple (36)] and smaller ascospores (6–7.5 × 3–3.5 µm vs. 9.5–13 × 4.5–6.6 µm with smooth perispore (vs. with conspicuous coil-like ornamentation) (Ju & Rogers 1996).
In surface coloration and ascospore size, H. populi is similar to H. lividocolor. However, the latter differs in having taller perithecia (0.5–1.3 mm high vs. 0.40–0.64 mm high), larger asci (135–160 µm total length vs. 104–125 µm total length), and Dark Livid (80) KOH-extractable pigments [vs. Umber (9) to Dark Purple (36)] (Ju & Rogers 1996). Other closely related species, H. lividipigmentum and H. hainanense, differ in having effused–pulvinate stromata and Dark Livid (80), Pale Vinaceous (85) to Livid Vinaceous (83) and Vinaceous Purple (101) KOH-extractable pigments. Moreover, H. hainanense has light-brown to brown, smaller ascospore 6.1–9.6 × 3.2–5 µm and H. lividipigmentum has reddish brown stromatal granules (Ju & Rogers 1996; Fournier et al. 2014; Ma et al. 2022). Based on these differences and phylogeny, we introduce Hypoxylon populi as a new species.
Hypoxylon pseudorubiginosum K. Habib, Y.H. Pi & Q.R. Li sp. nov.
MycoBank No.: MB859010; Fig. 13
Etymology: The epithet refers to the morphological and phylogenetically closely related Hypoxylon rubiginosum.
Material examined: China, Guizhou Province, Rongjiang County, Leigongshan Nature Reserve (26°21'49.43"N, 108°16'8.05"E), altitude: 1165 m, on dead wood, 28 Aug. 2020, Y. H. Pi, 2020LGS190 (GMB0295, holotype; GMBC0295 ex-type culture; KUN-HKAS 148621, isotype)
Teleomorph: Stromata on surface of dead wood, 1.5–2.5 cm long × 0.6–0.8 cm wide × 0.8–0.9 mm thick effused, surface Brown Vinaceous (84), Sepia (63), or Dark Brick (60); widely effused margins black, weakly carbonaceous, with orange-yellow granules beneath surface and between perithecia, the tissue below the perithecial layer conspicuous, with 10% KOH extractable pigments Luteous (12) to Umber (9), Dark Brick (86). Perithecia 0.35–0.68 mm wide × 0.53–0.69 mm high (x̄ = 0.53 × 0.60 mm, n = 20), spherical. Ostioles umbilicate, lower than the level of stromatal surface, surrounded by a ring of white substance. Asci 105–155 × 6.5–8.8 µm (av.= 130 × 8 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, the spore-bearing part 65–78 µm (x̄ = 71 µm, n = 30) long, with apical apparatus bluing in Melzer’s reagent, discoid, 2–2.8 µm broad × 0.8–1.4 µm high (x̄ = 2.4 × 1.2 µm, n = 20). Ascospores 9.5–11.6 × 4.5–6 µm (x̄ = 10.5 × 5 µm, n = 30), uniseriate, unicellular, ellipsoidal, inequilateral, with rounded ends, brown to blackish brown, with conspicuous straight germ slit covering the full spore length, perispore dehiscent in 10 % KOH, smooth, epispore smooth. Asexual morph: not formed.
Culture characteristics: Colonies on OA covering 4-5cm diam. after 2 weeks at 25 °C. Colonies surface white, cottony, azonate with entire margins; after 4 weeks, covering a 9 cm petri dish, become white grey, with yellow granules close to center; reverse black brown to hazel. No conidia were observed.
Additional material examined: China, Sichuan Province, Luzhou City, Huangjing Old Forest Nature Reserve, (28°6'6.2"N, 105°12'48.5"E), altitude 1000 m, on dead wood, 16 Aug. 2022, H.M. Hu, 2022HJLL25 (GMB4261).
Notes: Hypoxylon pseudorubiginosum is morphologically similar to H. rubiginosum, sharing comparable ascomatal and ascospore dimensions (Ju & Rogers 1996). Morphologically, H. pseudorubiginosum differs from H. rubiginosum (as described by Ju & Rogers 1996) in its KOH-extractable pigments Luteous (12) to Umber (9) or Dark Brick (86), contrasting with the orange (7) to rust (39) pigments of H. rubiginosum. Additionally, H. pseudorubiginosum possesses shorter asci (105–155 µm total length vs. 130–180 µm in H. rubiginosum). These distinctions are further corroborated by Pourmoghaddam et al. (2020), who reported H. rubiginosum yield Orange (7) to Scarlet (5) KOH pigments and asci stipes up to 180 µm long, further separating it from H. pseudorubiginosum. In the phylogram (Fig. 1), both species also showed deviating positions.
Hypoxylon pseudorubiginosum also shows superficial resemblance to H. petriniae due to shared vinaceous-brown stromata with black margins and ascospores of similar size. However, H. petriniae is distinguished by smaller ascomata (0.25–0.38 mm diam × 0.25–0.50 mm high vs. 0.35–0.68 mm diam × 0.53–0.69 mm high in H. pseudorubiginosum). Molecular data further confirm their distinction, with H. pseudorubiginosum exhibiting substantial genetic divergence from both H. rubiginosum and H. petriniae. Phylogenetically, it is distantly related, represent high molecular variation. Based on these differences and phylogenetic analysis, we introduce H. pseudorubiginosum as a new species.
Hypoxylon qiongense K. Habib, Y.H. Pi & Q.R. Li sp. nov.
MycoBank No.: MB859011; Fig. 14
Etymology: The specific epithet refers to Qiong, the traditional name for Hainan Province, the type locality.
Material examined: China, Hainan Province, Qiongzhong County, Limushan Nature Reserve (19°12'28.95"N, 109°50'21.74"E), altitude 209 m, on dead wood, 12 Nov. 2020, Y.H. Pi, 2020QZ45 (GMB4229, holotype; GMBC0449, ex-type culture; KUN-HKAS 148622, isotype).
Teleomorph: Stromata on surface of dead wood, 0.8–6 cm long × 0.3–3 cm wide × 0.7–0.95 mm thick (x̄ = 4 cm × 1.6 cm × 0.8 mm, n = 10), glomerate, coalescent into larger effused-pulvinate stromata, with conspicuous to inconspicuous perithecial mounds, noncarbonaceous, surface Blood Red (3) to Brown Vinaceous (84), with KOH-extractable pigments Fawn (87) to Sepia (63) after a while; tissue between the perithecia black, tissue below the perithecial layer conspicuous, blackish brown. Perithecia 0.46–0.64 mm wide × 0.61–0.94 mm high (x̄ = 0.55 × 0.82 mm, n = 20), subglobose or tubular. Ostioles umbilicate, lower than the surface, inconspicuous. Asci 90–106 × 5–6.5 µm (x̄ = 97 × 5.8 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, the spore-bearing part 45–60 µm (x̄ = 51 µm, n = 30) long, with apical apparatus bluing in Melzer’s reagent, discoid, 1.2–1.6 × 0.6–0.9 µm (x̄ = 1.5 × 0.8 µm, n = 20). Ascospores 6–8 × 3–3.5 µm (x̄ = 7 × 3 µm, n = 30), uniseriate, unicellular, ellipsoid inequilateral to equilateral, with rounded ends, light brown to brown, with an inconspicuous germ slit of spore length; perispore indehiscent in 10% KOH; epispore smooth. Asexual morph: not formed.
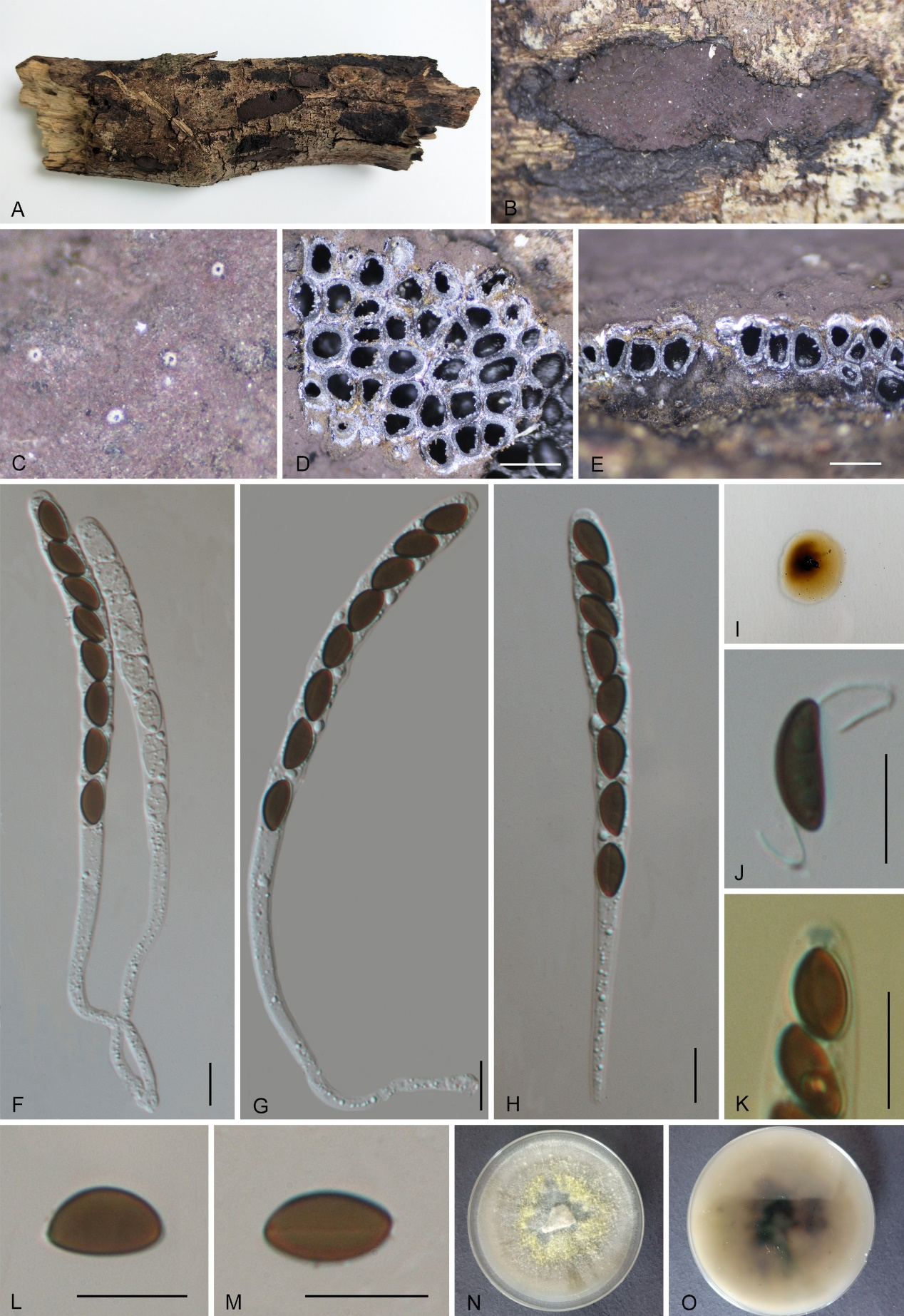
Culture characteristics: Colonies grown on OA at 25 °C for 3 weeks reached a diameter of 5 cm. Colonies surface white, felty, cottony, thin; reverse white. Not sporulating on OA nor on PDA.
Additional material examined: China, Hainan Province, Wuzhishan City, Atuoling Forest Park (18°48'26.79"N, 109°30'55.47"E), altitude 430 m, on dead wood, 15 Nov. 2020, Y.H. Pi and Q.R. Li, 2020ATL16-1 (GMB4265). China, Hainan Province, Wuzhishan City, Atuoling Forest Park (18°48'28.31"N, 109°30'58.63"E), altitude 571 m, on dead branches, 18 Nov. 2020, Y.H. Pi and Q.R. Li, 2020ATL306 (GMB0449)
Notes: Phylogenetically (Fig. 1) and morphologically, Hypoxylon qiongense is closely related to H. pulicicidum, sharing similar surface coloration, indehiscent perispore, and similarly sized asci. However, H. pulicicidum differs by its slightly larger ascospores (7.5–9.4 × 3.2–4.2 µm vs. 6–8 × 3–3.5 µm) and the production of Olivaceous Buff (89) pigments in 10% KOH [vs. Fawn (87) to Sepia (63)] (Bills et al. 2012). Sequence comparison also supports this differentiation, pairwise sequence identity between H. qiongense and H. pulicicidum (MUCL 49879) was 86.4% for ITS, 98.3% for LSU, and 95.5% for tub2.
Morphologically, Hypoxylon qiongense also resembles H. investiens, share effused-pulvinate stromata with a brown surface, an indehiscent perispore, and similarly sized asci. However, H. investiens differs in having tubular to long tubular perithecia, (vs. spherical in H. qiongense) and yield Greenish Yellow (16), Dull Green (70), or Dark Green (21) stromatal KOH-extractable pigments.
In perithecial, ascus, and ascospore size, Hypoxylon qiongense resembles H. hinnuleum and is also phylogenetically closely related. However, H. hinnuleum can be easily distinguished by its stromatal coloration, which ranges from Dark Vinaceous (82) to Sepia (63), becoming black at maturity, presence of orange-brown granules immediately beneath the surface and between perithecia (vs. black in H. qiongense) and yield Dark Brick (86) KOH-extractable pigments (Sir et al. 2019).
Based on these morphological differences and phylogeny, we introduce Hypoxylon qiongense as a new species.
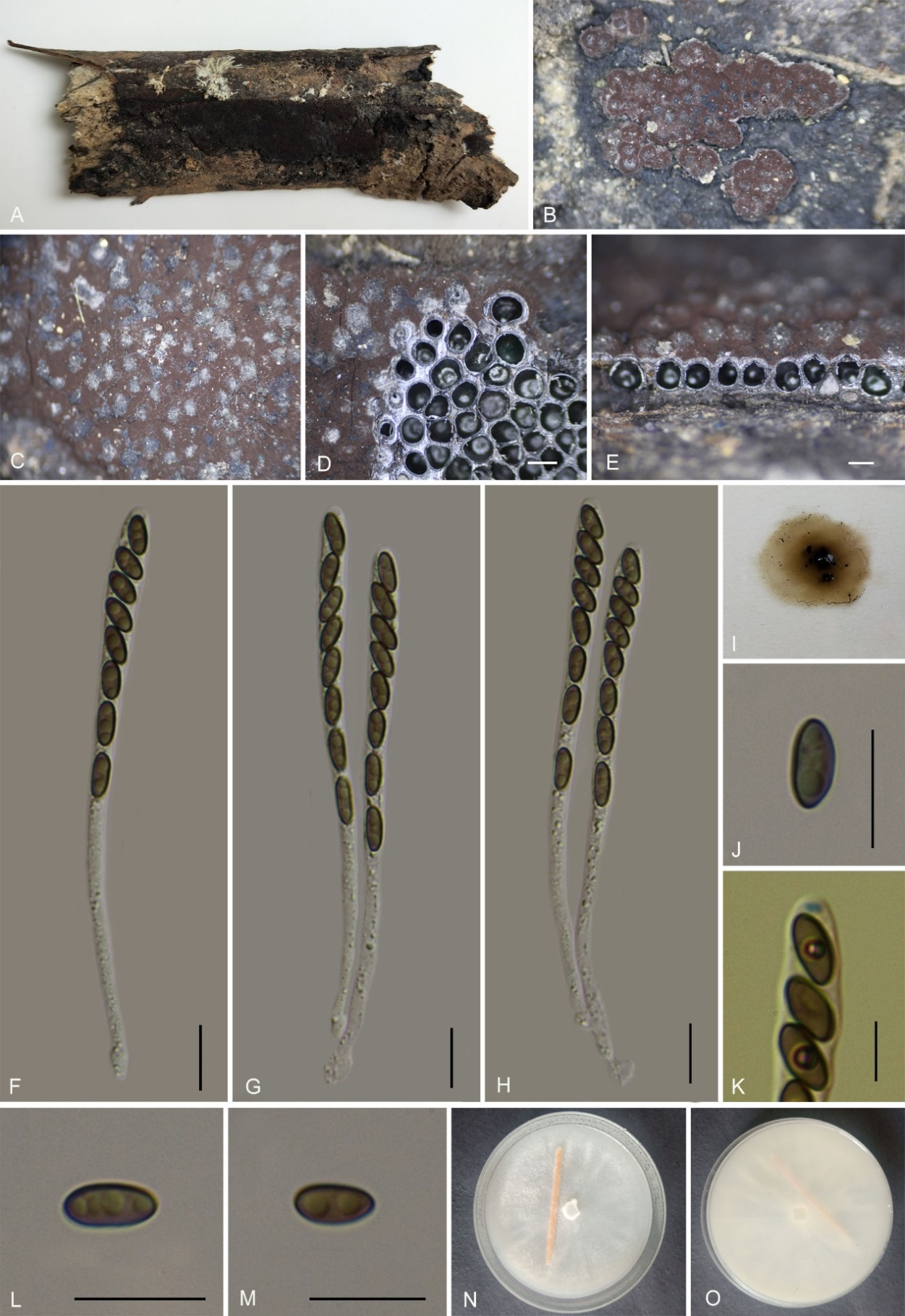
Hypoxylon rongjiangense K. Habib, Y.H. Pi & Q.R. Li sp. nov.
MycoBank No.: MB859004; Fig. 15
Etymology: The epithet refers to Rongjiang County, where the holotype was collected.
Material examined: China, Guizhou Province, Rongjiang County, Leigongshan Nature Reserve (26°22'53.08"N, 108°21'49.71"E), altitude: 881 m, on dead wood, 29 Aug. 2020, Y.H. Pi, 2019LGS52 (GMB0303, holotype; GMBC0303, ex-type living culture; KUN-HKAS 148623, isotype).
Teleomorph: Stromata on the surface of dead wood, 2.5–6 cm long × 1–2 cm wide × 0.5–0.55 mm thick (x̄ = 3 cm × 1.6 cm × 0.53 mm, n = 10), plane, effuse, surface Chestnut (40) or Dark Purple (36), orange granules beneath surface and between perithecia, tissue below the perithecial layer brownish, with 10% KOH extractable pigments Orange (7) Sienna (8) or umber (9). Perithecia 0.27–0.52 mm wide × 0.45–0.61 mm high (x̄ = 0.39 × 0.52 mm, n = 20), spherical. Ostioles lower than the level of stromatal surface, usually overlain with conspicuous white substance. Asci 80–110 × 6–9 µm (x̄ = 93 × 7 µm, n = 30), 8-spored, unitunicate, cylindrical, short-stipitate, spore-bearing part 60–76 µm (x̄ = 66 µm, n = 30) long, stipes 25–35 µm (x̄ = 28 µm, n = 30) long, with apical apparatus bluing in Melzer’s reagent, discoid 1.7–2 wide × 0.9–1.2 µm high (x̄ = 1.8 × 1 µm, n = 20). Ascospores 8.5–9.7 × 4–5.5 µm (x̄ = 9 × 5 µm, n = 30), uniseriate, unicellular, ellipsoidal, inequilateral, with broadly rounded ends, brown to dark brown, with conspicuous straight to slightly sigmoid germ slit covering the full spore length, perispore dehiscent in 10% KOH, smooth; epispore smooth. Asexual morph: not formed.
Culture characteristics: Colonies on OA covering 4–5 cm diam. after 3 weeks at 25 °C. Colonies surface white, cottony, azonate with entire margins; after 6 weeks, covering a 9 cm Petri dish, become brownish, appressed with entire margins; reverse grey brown to hazel. No conidia were observed.
Additional material examined: China, Guizhou Province, Xishui County, Danxia Valley of Xishui National Nature Reserve (18°48'36.44"N, 109°34'12.26"E), altitude 1094 m, on dead wood, 18 Nov. 2022, H.M. Hu, 2022DXG13 (GMB4714). China, Guizhou Province, Rongjiang County, Leigongshan Nature Reserve (26°22'53.22"N, 108°21'49.31"E), altitude 865 m, on dead wood, 29 Aug. 2020, Y.H. Pi, 2019LGS69 (GMB0304).
Notes: Morphologically, Hypoxylon rongjiangense is similar to H. zangii in having the same surface coloration, ascomata overlain with a conspicuous white substance, and yielding similar KOH-extractable pigments. However, H. zangii differs in having smaller ascomata (0.20–0.40 mm wide × 0.30–0.50 mm high vs. 0.27–0.52 mm wide × 0.45–0.61 mm high), larger asci (85–145 µm total length vs. 80–110 µm), and larger ascospores (10.9–14.6 × 4.8–6.4 µm vs. 8.5–9.7 × 4–5.5 µm) (Song et al. 2022).
In surface coloration and ascospore size, H. rongjiangense also resembles H. texense. However, H. texense can be distinguished by its KOH-extractable pigments Rust (39) to Dark Brick (86), and its larger ascomata (0.3–0.6 mm high × 0.28–0.5 mm diam) (Sir et al. 2019).
Hypoxylon rongjiangense shares similar KOH-extractable pigmentation with H. guilanense but the latter differs by having hemispherical to pulvinate stromata with Sienna (8), Umber (9) to Buff (45) surface colors, conspicuous perithecial mounds, and larger ascospores (12–15 × 5–6 µm) (Pourmoghaddam et al. 2020). There are sufficient nucleotide differences among the analyzed loci between H. rongjiangense and its closely related taxa, including H. zangii, H. texense, H. guilanense, H. canariense, and H. laschii. Both molecular and morphological evidence support the recognition of H. rongjiangense as a new species in the genus Hypoxylon.

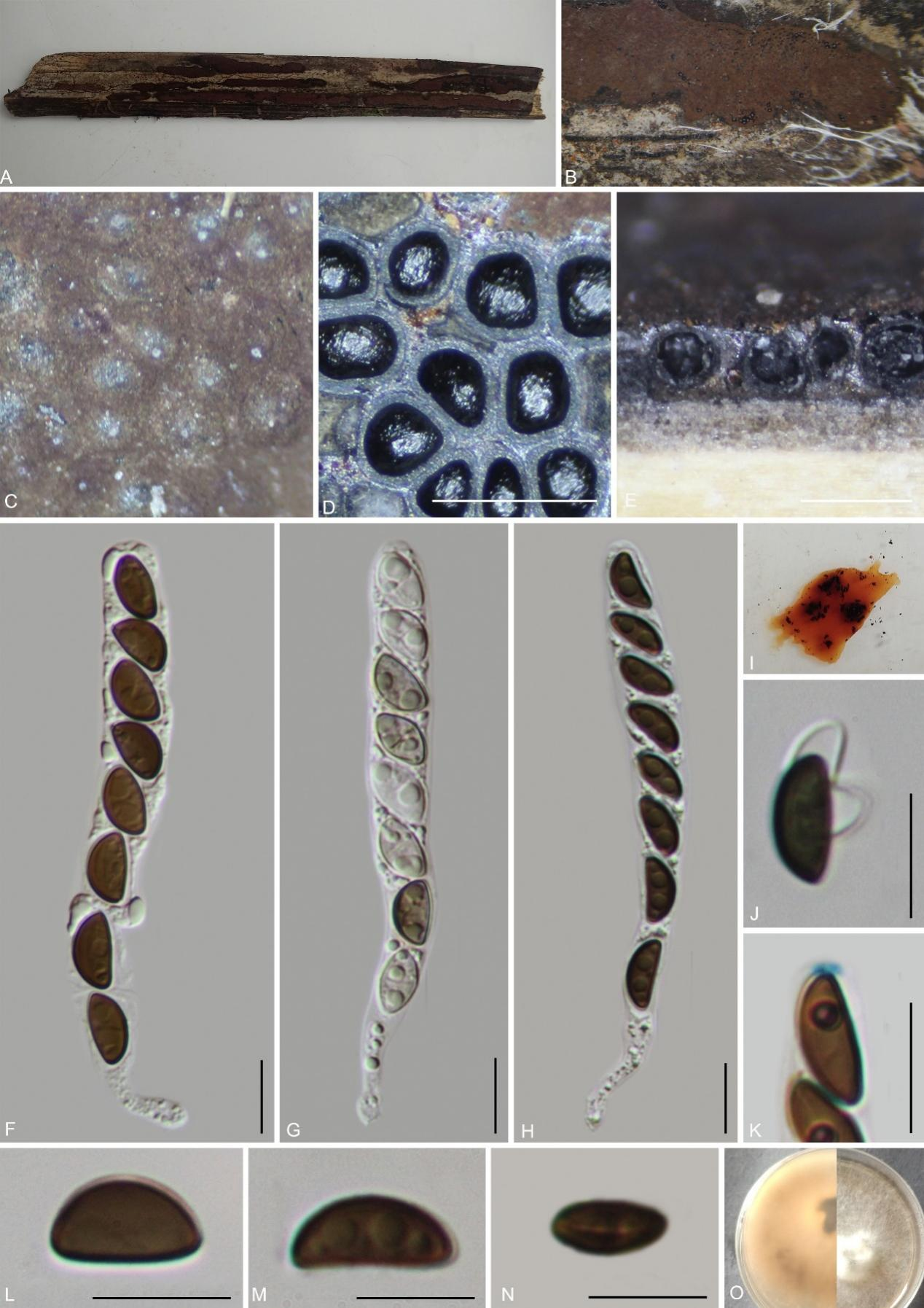
Hypoxylon sichuanense H.M. Hu, K. Habib & Q.R. Li sp. nov.
MycoBank No.: MB859012; Fig. 16
Etymology: The specific epithet “sichuanense” refers to the type locality Sichuan Province, from where the specimen was collected.
Material examined: China, Sichuan Province, Huangjing Old Forest Nature Reserve, (28°6'54.9"N, 105°14'2.9"E), altitude 800 m, on dead bamboo culm, 16 Aug. 2022, H.M. Hu, 2022HJLL36 (GMB4252, holotype; GMBC4252 ex-type; KUN-HKAS 148624, isotype)
Teleomorph: Stromata on surface of dead bamboo culm,1.2–6 cm long × 0.7–5 cm broad × 0.2–0.3 mm thick, effused or effused-pulvinate, confluent and forming larger elongate stromata, carbonaceous, surface Dark Vinaceous (82) or Sepia (63), yellowish-brown granules immediately beneath stromata, tissue black between perithecia, tissue below the perithecia inconspicuous, black, with 10% KOH extractable pigments Umber (9) to Rust (39). Perithecia 0.26–0.35 mm wide × 0.23–0.40 mm high (x̄ = 0.32 × 0.34 mm, n = 20), globose to subglobose, black. Ostioles slightly lower than the level of stromatal surface, without a disc. Asci 86–110 × 9–11.5 µm (x̄ = 98 × 10.5 µm, n = 30), 8-spored, unitunicate, cylindrical, spore-bearing part 68–77 µm long (x̄ = 71 µm, n = 30), with apical apparatus bluing in Melzer’s reagent, discoid, 2–3 × 1.5–2.2 µm (x̄ = 2.7 × 1.8 µm, n = 20). Ascospores 10–11.5 × 5–6 µm (x̄ = 10.5 × 5.5 µm, n = 30), uniseriate, unicellular, brown to dark brown, ellipsoid, inequilateral, with rounded ends, smooth, with an inconspicuous straight germ slit covering the full spore length; perispore dehiscent in 10% KOH, smooth; epispore smooth. Asexual morph: Not observed.
Culture characteristics: Colonies on OA covering 4 cm diam. after 3 weeks at 25 °C, Colonies surface white, felty to cottony, azonate; after 6 weeks covering a 9 cm petri dish, becoming greyish white, dense; reverse brownish yellow. No conidia were observed.
Additional material examined: China, Sichuan Province, Luzhou City, Huangjing Old Forest Nature Reserve (28.115037°N, 105.234028°E), altitude: 583 m, on dead bamboo culm, 20 Aug. 2022, H.M. Hu, 2022HJLL301 (GMB4711).
Notes: Morphologically and phylogenetically (Fig. 1), Hypoxylon sichuanense is closely related to H. pilgerianum, which has also been reported on bamboo. However, H. pilgerianum differs in having dull orange-brown or dull reddish-brown granules immediately beneath the surface and between the perithecia (vs. black between the perithecia), smaller asci (68–97 µm total length vs. 86–110 µm total length), and slightly longer, narrower ascospores with narrowly rounded ends (8.5–13.5 × 4–5.5 µm vs. 10–11.5 × 5–6 µm with rounded ends) (Ju & Rogers 1996).
Hypoxylon pilgerianum was originally described from Brazil on Chusquea sp., with ascospores 10–12 × 4–5 µm. Miller (1961), treated it as a synonym of H. rubiginosum. Later, Ju & Rogers (1996) reinstated the species based on collections from dead bamboo culms, reporting ascospores of 8.5–12(–13.5) × 4–5(–5.5) µm. Fournier et al. (2015) documented two collections from Martinique, designating them as H. cf. pilgerianum sp. 1 and sp. 2, with ascospores 7.6–10 × 3.4–4.4 µm and 10.3–12.8 × 4.9–6.7 µm, respectively. In terms of chemical profiles, Ju & Rogers (1996) reported KOH-extractable pigments of Ochreous (44), Honey (64), or Amber (47), whereas Fournier et al. (2015) observed a pigment Sienna (8) turning Greyish Sepia (106). In contrast, H. sichuanense yields pigments Umber (9) to Rust (39).
Other species reported on bamboo, including H. wuzhishanense, H. tibeticum, and H. pseudefendleri, share brownish stromata but all differ from H. sichuanense by the presence of yellowish-brown granules beneath the surface and between the perithecia (vs. black in H. sichuanense). Moreover, H. wuzhishanense has larger ascospores (9.5–14 × 5.4–6.7 µm) and an indehiscent perispore (Ma et al. 2022). Hypoxylon tibeticum has larger ascospores (9.8–13 × 5.1–6.9 µm). Hypoxylon pseudefendleri has larger stromata (0.50–0.85 mm diam × 0.35–0.50 mm high), and shorter asci (70–90.5 µm total length). There are no descriptions of its stromatal pigments in 10% KOH, and perispore detail, so these characteristics cannot be compared (Dai et al. 2017). Phylogenetic analyses also support the distinction of H. sichuanense. Hence, we introduce it as a new species.
Hypoxylon subjaklitschii H.M. Hu, K. Habib & Q.R. Li sp. nov.
MycoBank No.: MB859013; Fig. 17
Etymology: The epithet refers to the morphologically similar species, H. jaklitschii.
Material examined: China, Hainan Province, Anlong County (18°57'15.15"N, 109°03'45.21"E), altitude 1032 m, on dead wood, 22 July 2022, H.M. Hu, 2022BWL16 (GMB4241, holotype; GMBC4241, ex-type; KUN-HKAS 148625, isotype).
Teleomorph: Stromata on the surface of dead wood, 0.1–0.8 cm long × 0.1–0.4 cm wide × 0.5–1.0 mm thick (x̄ = 0.38 cm × 0.22 cm × 0.89 mm, n = 10), pulvinate, glomerate, solitary to aggregated, spherical, united by thin stromatal tissue, non-carbonaceous, surface Sepia (63), Dark Brick (66) to black, black beneath surface and between perithecia, tissue below the perithecial layer inconspicuous, black, with KOH-extractable pigments Umber (9) or Brick (59). Perithecia 0.57–0.78 mm wide × 0.60–0.86 mm high (x̄ = 0.67 × 0.71 mm, n = 20), spherical. Ostioles slightly higher than stromatal surface. Asci 130–170 × 8–10.5 µm (x̄ = 148 × 8.9 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, the spore-bearing part 76–98 µm (x̄ = 88 µm, n = 30) long, apical apparatus not bluing in Melzer’s reagent. Ascospores 11–13.5 × 5.2–6.6 µm (x̄ = 12.2 × 6.2 µm, n = 30), uniseriate, unicellular, ellipsoidal, inequilateral to nearly equilateral, with rounded ends, brown to dark brown, with a sigmoid spore-length germ slit, perispore dehiscent in 10% KOH with inconspicuous ornamentation; epispore smooth. Asexual morph: not formed.
Culture characteristics: Colonies on OA covering 3 cm diam. after 2 weeks at 25 °C, Colonies surface white, felty, azonate, margin thin; after 6 weeks covering a 9 cm petri dish, becoming yellow-white, thick at center; reverse brown. No conidia were observed.
Additional material examined: China, Hainan Province,
Anlong County (18°57'12.48"N, 109°03'42.59"E), altitude 985 m, on dead wood, 25 July 2022 H.M. Hu, 2022 BWL316 (GMB4722).

Notes: Morphologically, Hypoxylon subjaklitschii resembles H. jaklitschii in having the same surface coloration and stromatal KOH-extractable pigments. Phylogenetically (Fig. 1), they also appear as sister species. However, H. jaklitschii differs by having orange-brown to dark brown granules immediately beneath the surface and between the perithecia (vs. black), amyloid ascal apical apparatus (vs. non-amyloid), smaller perithecia (0.3–0.4 mm wide vs. 0.57–0.78 mm wide), umbilicate ostioles lower than the stromatal surface, and smaller ascospores [9.5–11.5(−12) × 4–5.5 µm vs. 11–13.5 × 5.2–6.6 µm] (Kuhnert et al. 2015).
In having glomerate stromata and similar KOH-extractable pigments, H. subjaklitschii also resembles H. lenormandii. However, H. lenormandii is morphologically distinct with its Grayish Sepia (102), Fuscous (103), or Brown Vinaceous (84) surface coloration [vs. Sepia (63), Dark Brick (66) to black], smaller perithecia (0.3–0.5 mm diam.), and larger ascospores (11–17 × 4–6.5 µm) (Kuhnert et al. 2015).
Phylogenetically, H. subjaklitschii is also closely related to Hypoxylon mangrovei and H. teeravasati. Hypoxylon mangrovei can be distinguished by its larger ascospores (12–14.5 × 5–6.5 µm) and orange KOH-extractable pigments (Dayarathne et al. 2020). Hypoxylon teeravasati differs by having a burnt sienna stromatal surface, amyloid ascal apical apparatus and eye brown, KOH-extractable pigments (Phookamsak et al. 2019).
Jackrogersella maolanensis K. Habib, Y.H. Pi & Q.R. Li sp. nov.
MycoBank No.: MB859014; Fig. 18
Etymology: The epithet refers to Maolan National Nature Reserve, where the holotype was collected.
Material examined: China, Guizhou Province, Maolan National Nature Reserve, (25°10'54.6"N, 108°8'31.4"E), altitude 586 m, on dead branch of an unidentified plant, 18 July 2022, H.M. Hu, 2022MLB32 (GMB0115, holotype; GMBC0115, ex-type; KUN-HKAS 148626, isotype).
Teleomorph: Stromata on the surface of dead wood of unidentified plant, 0.1–2.7 cm length, 0.4–1.2 cm wide, and 0.2–0.8 mm high, pulvinate, solitary to confluent, spherical, with individual perithecial mounds prominent and occasionally approaching a rosellinioid appearance, united by thin stromatal tissue, surface blackish, carbonaceous, tissue beneath perithecial layer inconspicuous, black; with KOH extractable pigments Dark Brick (60), Vinaceous Grey (116) pigments in 10% KOH. Perithecia 0.54–0.71 mm wide × 0.62–0.87 mm high (x̄ = 0.60 × 0.71 mm, n = 20), spherical to obovoid. Ostioles papillate, black. Asci 100–135 × 3–4 µm (x̄ = 122 × 4 µm, n = 30), 8-spored, unitunicate, cylindrical, long-stipitate, the spore-bearing part 40–52 µm (x̄ = 48 µm, n = 30) long, apical apparatus bluing in Melzer’s reagent, discoid, 1.6–2.1 × 0.6–1 µm (x̄ = 1.8 × 0.8 µm, n = 20). Ascospores 5.5–7.7 × 2.5–3.5 µm (x̄ = 6.4 × 3.1 µm, n = 30), uniseriate, unicellular, ellipsoidal, inequilateral, with rounded ends, light brown, with an inconspicuous germ slit; perispore dehiscent in 10% KOH, smooth, with a thickening on the convex side, epispore smooth. Asexual morph: not formed.
Culture characteristics: Colonies on OA covering 2 cm diam. after 2 weeks at 25 °C, Colonies surface white, thin felty, fluffy azonate; after 6 weeks covering a 9 cm petri dish, becoming brownish white, dense; reverse greyish brown. No conidia were observed.
Additional material examined: China, Guizhou Province, Kaili City, Leigongshan National Nature Reserve, (26°24'44.7"N, 108°12'4.5"E), altitude 960 m, on dead branch of an unidentified plant, 16 Aug. 2022, H.M. Hu, 2022LGS5 (GMB0114).
Notes: Phylogenetically (Fig. 1), Jackrogersella maolanensis is closely related to J. ilanensis, with which it shares similar surface coloration, and ascospore size. However, J. ilanensis can be distinguished by its effused pulvinate stromata, ostiole encircled by a truncatum-type disc overlain with a white substance (vs. lack), and shorter asci (95–110 µm total length vs. 100–135 µm total length) (Ju & Rogers 1996).
Jackrogersella maolanensis stromata could be confused with Jackrogersella minutella. However, J. minutella differs in having inconspicuous perithecial mounds, smaller perithecia (0.40–0.50 mm wide vs. 0.54–0.71 mm wide), an inamyloid apical ring, and larger ascospores (6–9 × 3–4 µm vs. 5.5–7.7 × 2.5–3.5 µm) with a straight germ slit shorter than the spore length (as H. cohaerens var. microsporum; Ju & Rogers 1996; Wendt et al. 2018).
Species of the genus that lack molecular data, J. gombakensis and J. nothofagi can be easily distinguished by surface coloration and ascospore size. Jackrogersella gombakensis has a dark reddish-brown stromatal surface and larger ascospores (13–15 × 5–6 μm). Jackrogersella nothofagi has effused-pulvinate stromata with inconspicuous to planar perithecial mounds, a dark vinaceous to brown vinaceous surface, and larger ascospores 7–9.9 × 3.5–5 μm (Ju & Rogers 1996; Wendt et al. 2018).
Mitochondrial Genome
Characterization of the fifty Hypoxylaceae mitogenomes
The mitochondrial genomes of the 50 Hypoxylaceae strains represent four genera Annulohypoxylon, Hypomontagnella, Hypoxylon, and Jackrogersella. All mitogenomes are circular DNA molecules. Annotated mitochondrial diagrams of three representative species are shown in Figure 19. The remaining circular diagrams are in Supplementary Figure 7. A detailed overview of the mitochondrial genome composition across 50 strains within Hypoxylaceae is provided in Supplementary Tables 10 and 11.
The genomes are composed of circular DNA molecules with highly variable sizes across genera. In Hypomontagnella, genome sizes range from 27,637 bp (H. vinacea GMB0247) to 47,204 bp (H. yinggelingensis GMB4715). Hypoxylon shows the widest variation, ranging from 26,809 bp (H. lancangense GMB4249) to 197,900 bp (Hypoxylon sp. GMB0308). Annulohypoxylon exhibits comparatively larger genomes, ranging from 130,895 bp (A. baishaense GMB4213) to 251,279 bp (A. viridipigmentum GMB0225). Jackrogersella maolanensis (GMB0115) has a genome size of 157,709 bp, which falls between the upper range of Hypoxylon and the lower range of Annulohypoxylon (Supplementary Fig. 7).
Protein coding genes and introns of the mitochondrial genomes
All 50 mitochondrial genomes of Hypoxylaceae contain 14 conserved core protein-coding genes (PCGs), including atp6, atp8, atp9, cob, cox1, cox2, cox3, nad1, nad2, nad3, nad4, nad4L, nad5, nad6 (Fig. 19, Supplementary Table 11). Among these, nad5 was the longest PCG, with an average length of 2,416.4 bp, while atp8 was the shortest, averaging 153.6 bp. The atp8 gene was the shortest PCG across all 50 mitochondrial genomes. Its length was 153 bp in all strains except Annulohypoxylon subyungense GMB0226 and A. pseudoalbidiscum GMB0235, which showed 168 bp (Supplementary Table 11).
Gene length was most variable for nad3, ranging from 225 bp (Hypoxylon sp. GMB0308) to 2,631 bp (Jackrogersella maolanensis GMB0115). The gene cox3 showed the second-highest variation, spanning 483 bp to 1,872 bp across species. Of the 14 core PCGs, only atp9 was invariant in length. The gene length of nad1 in Annulohypoxylon primevalense GMB0240 (1,110 bp vs. 1,107 bp in others) and nad4L in Hypoxylon sp. GMB4719 (237 bp vs. 270 bp in others) deviated from those in the other 49 strains (Supplementary Table 11).
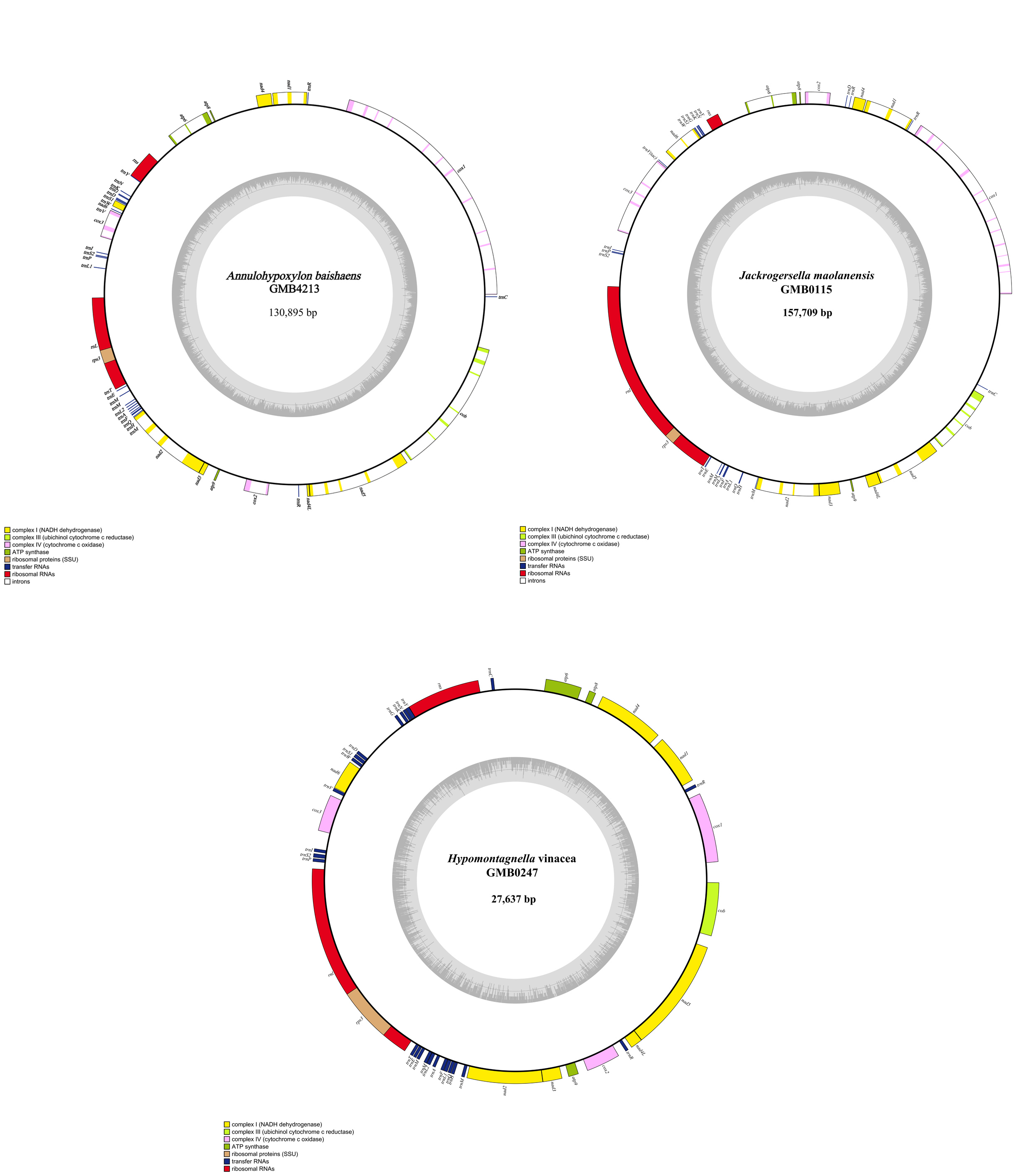
Intron content varied significantly across the family, ranging from a complete absence in Hypomontagnella vinacea GMB0247 and Hypoxylon lancangense GMB0284 and GMB4249, to a maximum of 68 introns (with a cumulative intronic length of 64,743 bp) in Annulohypoxylon linjiangense GMB4216, which accounts for 45% of its total genome. Among the 14 core protein-coding mitochondrial genes (cox1, cox2, cox3, cob, atp6, atp8, atp9, nad1, nad2, nad3, nad4, nad4L, nad5, nad6), introns were detected in all genes except atp9, nad1, atp8 and nad4L (Supplementary Tables 10, 11). The nad3 gene showed the highest intron diversity, with variable intron counts (225 bp in Hypoxylon sp. GMB0308 vs. 2631 bp in Jackrogersella maolanensis GMB0115), suggesting frequent intron gain or loss events.
Annulohypoxylon microdiscum GMB0234 has a short cox1 (1,284 bp) and intron size of 81,032 bp, suggesting minimal intronic contribution to cox1. Annulohypoxylon pseudoalbidiscum GMB0235 has an exceptionally long cox3 (1,872 bp) and intron size of 59,837 bp, implying a large intron within cox3.
Intron-rich species included Annulohypoxylon linjiangense GMB4216 (68 introns, 64,743 bp), A. viridipigmentum GMB0225 (50 introns, 120,275 bp), A. yunnanense GMB0462 (46 introns, 104,603 bp), Hypoxylon duranii GMB0265 (47 introns, 96,542 bp), and Hypoxylon sp. GMB0308 (43 introns, 110,871 bp).
Intergenic regions and rRNA and tRNA
The intergenic regions also showed high variation, ranging from 4,781 bp in Hypomontagnella vinacea GMB0247 (~17.3% of its 27,637 bp genome) to 62,450 bp in Hypoxylon sp. GMB0308 (~31.6% of its 197,900 bp genome). All strains encoded 25–35 tRNA genes (average ~26) and consistently 2 rRNA genes, with tRNA sizes ranging from 1,840 bp (Annulohypoxylon subyungense GMB0226) to 2,565 bp (Hypoxylon sp. GMB0297), and rRNA sizes ranging from 5,214 bp (Hypoxylon qiongense GMB4265) to 33,434 bp (Annulohypoxylon areolatum GMB4224) (Supplementary Table 10). This variability highlights the structural plasticity of Hypoxylaceae mitogenomes, indicating conserved rRNA gene numbers but significant divergence in intergenic regions and tRNA gene composition, likely reflecting evolutionary adaptations to diverse ecological niches, include expansions in non-coding regions for regulatory flexibility or tRNA/rRNA optimization to enhance metabolic efficiency.
Comparison of genes and similar species based on mitochondrial genomes
All 50 examined specimens possess the same set of 14 conserved mitochondrial protein-coding genes, indicating no variation in gene presence or absence across species. However, their mitochondrial genome sizes vary significantly, ranging from 26,809 bp (Hypoxylon lancangense GMB4249) to 251,279 bp (Annulohypoxylon viridipigmentum GMB0225). The lengths of the protein-coding genes show varying degrees of conservation and variability, which forms the basis of the correlation between genes and species.
Detailed analysis of gene lengths showed that some genes nad1, atp8, atp9, and nad4L, are highly conserved in length across species, suggesting strong functional constraints. For example, nad1 gene of nearly all species has a length of 1,107 bp, with only Annulohypoxylon primevalense GMB0240 at 1,110 bp (a difference of just 3 bp). Similarly, atp8 is 153 bp in most species, with only two exceptions (Annulohypoxylon pseudoalbidiscum GMB0235 and Annulohypoxylon subyungense GMB0226) at 168 bp. The gene atp9 is uniformly 225 bp across all species, showing no variation, representing the most conserved gene in the dataset.
In contrast, genes like nad3 (225–2,631 bp) show extreme variability, driven largely by species-specific intron content. The gene length variation ranges overlap significantly across genera, and no consistent genus-specific pattern emerges, indicating that gene length variation is more species-specific than genus-specific.
The observed variability in gene lengths, particularly for genes like cox1, cob, and nad3, is likely influenced by the presence and size of introns within these genes. For example, Annulohypoxylon microdiscum GMB0234 has a short cox1 (1284 bp) despite a large total intron content (81,032 bp), suggesting that cox1 itself contains few or no introns and that most intronic sequence is located elsewhere in the mitogenome. Hypomontagnella yinggelingensis GMB4715 has a longer cox1 (1,833 bp) with a total intron size of 6,740 bp, possibly indicating the presence of introns within cox1. Similarly, Annulohypoxylon pseudoalbidiscum GMB0235 has an exceptionally long cox3 (1872 bp) with a total intron size of 59,837 bp, implying the presence of a large intron within cox3.
Genes like nad1, atp8, atp9, and nad4L are highly conserved in length, showing minimal variation across species. Genes like cox1, cob, nad4, and especially nad3 display greater length variability, likely due to species-specific differences in intronic content within these genes. This variability does not align with genus-level patterns but reflects individual species differences, possibly driven by evolutionary changes in intron presence or size. Significant mitochondrial genome size differences (26,809 to 251,279 bp), gene lengths remain relatively stable, indicating that genome size variation primarily attributed to non-coding regions (intergenic and intronic) rather than coding sequences.
The genus Hypomontagnella exemplifies mitochondrial diversity within a taxonomic group. Hypomontagnella vinacea (27,637 bp, no introns) and H. yinggelingensis (47,204 bp, two introns) differ significantly in genome structure, supporting mitochondrial features as potential supplementary barcodes. Hypoxylon lancangense strains GMB0284 and GMB4249 display highly similar mitochondrial features. Their genome sizes (~26,813 bp and 26,809 bp), GC content (26.5%), intron absence, and conserved protein-coding gene content are nearly identical, indicating strong intraspecific consistency and genomic stability within this species.
Annulohypoxylon microdiscum strains GMB0233 and GMB0234 are morphologically indistinguishable. However, their mitochondrial genomes show much differences in size (171,360 bp vs. 162,152 bp), intron number (39 vs. 36), and intergenic region lengths (51,305 bp vs. 41,635 bp). These genomic differences suggest the possibility of cryptic speciation or divergence at the genomic level. Due to the absence of clear morphological distinctions, lack of a type strain, and a lack of detailed genomic analyses we tentatively identify both strains as Annulohypoxylon microdiscum. Refer Supplementary Tables 10 and 11 for a comparison of other species.
Mitochondrial genomes provide insights into evolutionary processes, however, their utility for species delimitation is constrained by overlapping length ranges across genera and insufficient functional gene analysis. Genome size variation (CV ~50%) far exceeds coding sequence variability (CV 0–15%), emphasizing that non-coding regions dominate mitochondrial evolution. A polyphasic approach combining mitogenomic data with nuclear markers (ITS, LSU, rpb2, tub2) and detailed morphological analyses will be essential to refine species boundaries and develop robust mitochondrial barcode markers. Pairwise comparisons of gene sequences between identical and closely related species are summarized in Supplementary Tables 1–7.
Evolutionary rates of mitochondrial genes
The Ka/Ks ratios for nine of the 14 core PCGs and rps3 gene were less than 1, indicating that these nine genes were subjected to purifying selection during evolution. However, the Ka/Ks values of the atp8, cox1, nad2, nad3, and nad5 genes were >1 between some Hypoxylaceae species, including members of the genera Annulohypoxylon, Hypoxylon, Hypomontagnella and Jackrogersella indicating that these genes may be under strong positive selection pressure in certain lineages within Hypoxylaceae.
Analysis of Codon Preferences
A comparative analysis of codon usage trends in 50 Hypoxylaceae mitogenomes were performed (Fig. 21). The codons AGA (arginine) and UUA (leucine) were the most frequently utilized across the 50 Hypoxylaceae strains. In addition, the 50 Hypoxylaceae mitogenomes constantly exhibited a codon usage bias for certain amino acids, with UCG (Serine), ACU (Threonine), GGU (Glycine), GUC (Valine), AGU (Serine), and CCU (Proline) being strongly overrepresented (RSCU > 1).
Translation was primarily initiated with the start codon ATG, while UAA was the most common stop codon. Variation between preferred and avoided codons was observed but relatively modest, with UUA used approximately 120 times more than CUC in most strains. This pattern, in which A/U-ending codons are favored while the full set of synonymous codons is retained, is typical of AT-rich mitochondrial genomes reflects the high adenine (A) and thymine (T) content of their coding sequences.
This consistent codon preference observed among Hypoxylaceae strains suggests a family-wide pattern. However, significant variations were noted in the start and stop codons across different strains (Supplementary Fig. 5). The pace of protein translation and impacts energy expenditure in most organelles is stimulated by codon usage. Codon usage analysis reveals substantial bias in synonymous codon selection within the mitochondrial coding sequences.
Evolutionary rates of core genes
The lengths and GC contents of the 14 core PCGs varied among the 50 Hypoxylaceae mitogenomes and revealed distinct evolutionary patterns shaped by gene-specific constraints and phylogenetic relationships.
The average GC content of atp9 (35.4%) and cox1 (32.5%) exhibited consistently highest means, while nad6 (21.5%) and atp8 (20.8%) exhibited the lowest means of GC content, indicating relative conservation and structural constraints of these genes across the family. In a genus-level comparison, Annulohypoxylon strains exhibited slightly higher overall GC content (range 29.2–32.2%) than Hypoxylon and Hypomontagnella (range 26.4–31.6%), while Jackrogersella maolanensis had a GC content of 31.6%, clusters closer to Annulohypoxylon, revealing a close phylogenetic affinity. Among the 50 Hypoxylaceae mitogenomes, the Annulohypoxylon pseudoalbidiscum exhibited the highest rps3 GC content (34.8%), suggesting lineage-specific adaptation (Fig. 22).
The gene lengths of the 14 PCGs and rps3 revealed a core pattern of high evolutionary conservation and genus- and species-specific dynamic variation. The gene lengths of atp8 (153 bp), atp6 (795 bp), atp9 (225 bp), nad4L (270 bp), and nad1 (1,107 bp) were highly stable and showed minimal to no variation (Fig. 23), indicating strong functional constraints. The genus Hypoxylon exhibited the highest gene lengths overall compared to other genera. In addition, the nad3 gene exhibited massive size expansions in Jackrogersella maolanensis (GMB0115: 2,631 bp) and several Hypoxylon strains (GMB0298: 2,595 bp; GMB4252: 2,574 bp; GMB4265 and GMB4229: 2,502 bp; GMB0270: 2,487 bp) compared to its ordinary ~420 bp baseline. The GC skew for the 14 core PCGs was positive for all genes except atp8, which was consistently negative, revealing a unique evolutionary or mutational pressure on this gene (Fig. 24). The atp9 and rps3 exhibited the highest positive GC skews followed by nad3. Annulohypoxylon species exhibited higher and more homogenous GC skew values compared to other genera. In addition, higher variability or inverted GC skew was observed in atp6 and nad3 in Hypoxylon species. The AT skews across the 14 core PCGs were all negative across the 50 Hypoxylaceae mitogenomes. However, the rps3 gene exhibited consistent positive AT skew across all genera, revealing a unique evolutionary trajectory (Fig. 25). Compared to other genera, Annulohypoxylon species exhibited moderate negative AT skews for core genes like cox1 and nad1, indicating greater A/T compositional stability.
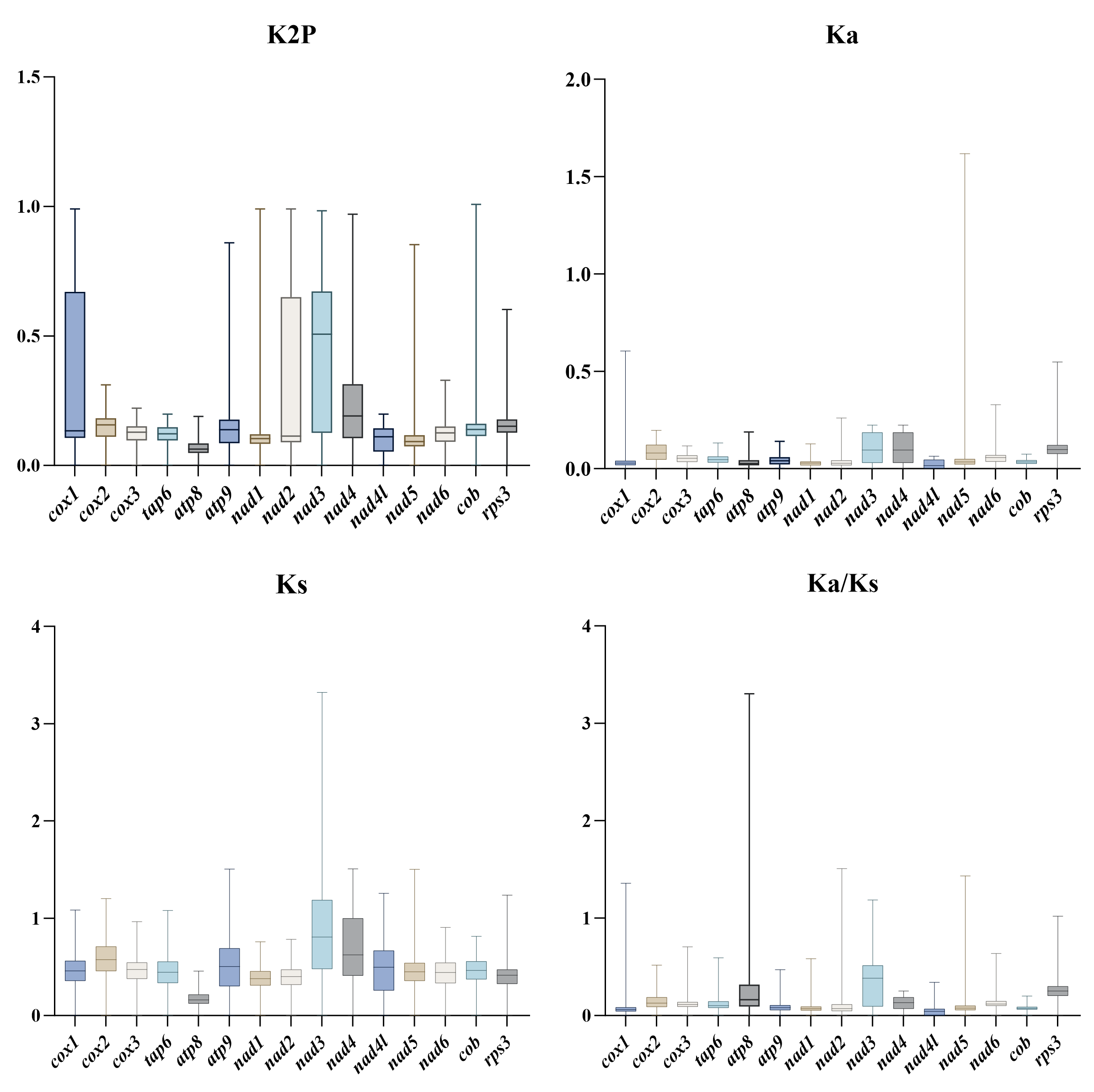
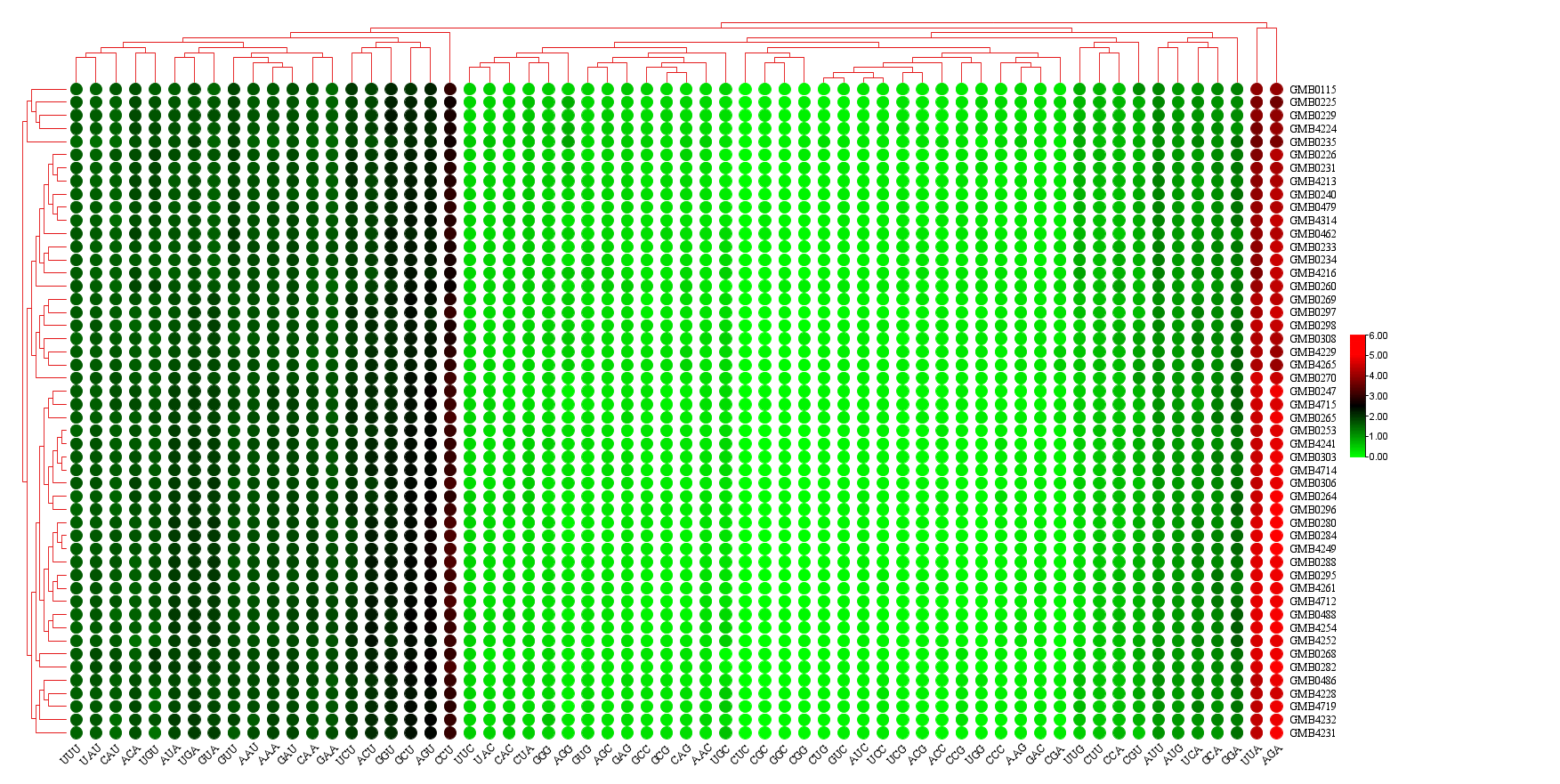
Gene arrangement
The order of mitogenome genes in the 50 Hypoxylaceae specimens revealed that 21 mitogenomes had the most conserved and identical gene order (cox1, trnR, nad1, nad4, atp8, atp8, atp6, rns, trnYNKGDSW, nad6, trnLV, cox3, trnISP, rnl, rsp3, trnTEMMLAFQHM, nad2, nad3, atp9, cox2, trnR, nad4L, nad5, cob and trnC). Most of the tRNAs, such as trnR, trnV, trnISSWP, trnYNKGDSW, trnYNKGW, trnYNKGGDW, trnISP, and trnW underwent translocation and duplications across the mitogenomes. (Fig. 26). The region between atp8 and rns shows high variation, in some mitogenomes there is no intervening gene, while others contain short tRNAs annotated as I, E, C, CR, L, RI, or R (Fig. 26).
The cox2 gene in Jackrogersella maolanensis (GMB0115) underwent a genomic rearrangement, moving from a position downstream of atp9 to a location upstream of atp8 . Combined with phylogenetic analysis (Fig. 2), it is possible that this cox2 gene rearrangement event was a critical factor in the formation of the Jackrogersella clade. Additional mitogenome data from other Jackrogersella species will be required to further validate this inference. The tRNA sequences following the rns locus also exhibit distinct characteristics compared to those of other species. The differences in tRNA types and arrangements between Annulohypoxylon and other related genera are evident in the tRNA sequences following the rns genes. Moreover, the tRNA sequence YNKG is typically conserved in Hypoxylon and Hypomontagnella, whereas it can exhibit variations in Annulohypoxylon.. Additionally, there are significant differences in tRNA types and arrangements among various branches, which may play a role in speciation. The tRNA sequence TEMMLAFLQHM following the rps3 gene was largely conserved, with minor variations detected in strains GMB0488, GMB0308, GMB0298, and GMB0297 (Fig. 26).
Mitogenome Composition Analysis
Comparative genomic analyses of the mitogenomes of Hypoxylaceae species exhibited genetic variations and rearrangements (Fig. 27). The size and location of homologous regions varied greatly among the 50 strains, even among species within the same genus. Variations in both the number and size of homologous regions across species and genera reflect the evolutionary and functional diversity within Hypoxylaceae. Annulohypoxylon viridipigmentum GMB0225 exhibited the highest number of homologous regions (39 regions) and the largest cumulative size, whereas A. baishaense GMB4213 exhibited a smaller size and the lowest number of homologous regions (35 regions) within the genus Annulohypoxylon. Similarly, Hypoxylon sp. GMB0308 exhibited the highest number of homologous regions (25) and the largest size, while H. lancangense GMB0284 and H. lancangense GMB4249 exhibited the smallest sizes, each with only 8 homologous regions within the genus Hypoxylon. In the genus Hypomontagnella, Hy. yinggelingensis GMB4715 exhibited larger genome size with 12 homologous regions compared to Hy. vinacea GMB0247 which exhibited the smallest size and 11 homologous regions. With only one sampled species in the genus Jackrogersella, J. maolanensis GMB0115 exhibited 34 homologous regions and a comparatively large size compared to several species in other genera among the 50 Hypoxylaceae. Variations in homologous regions across different species may be associated with differences in intron content.
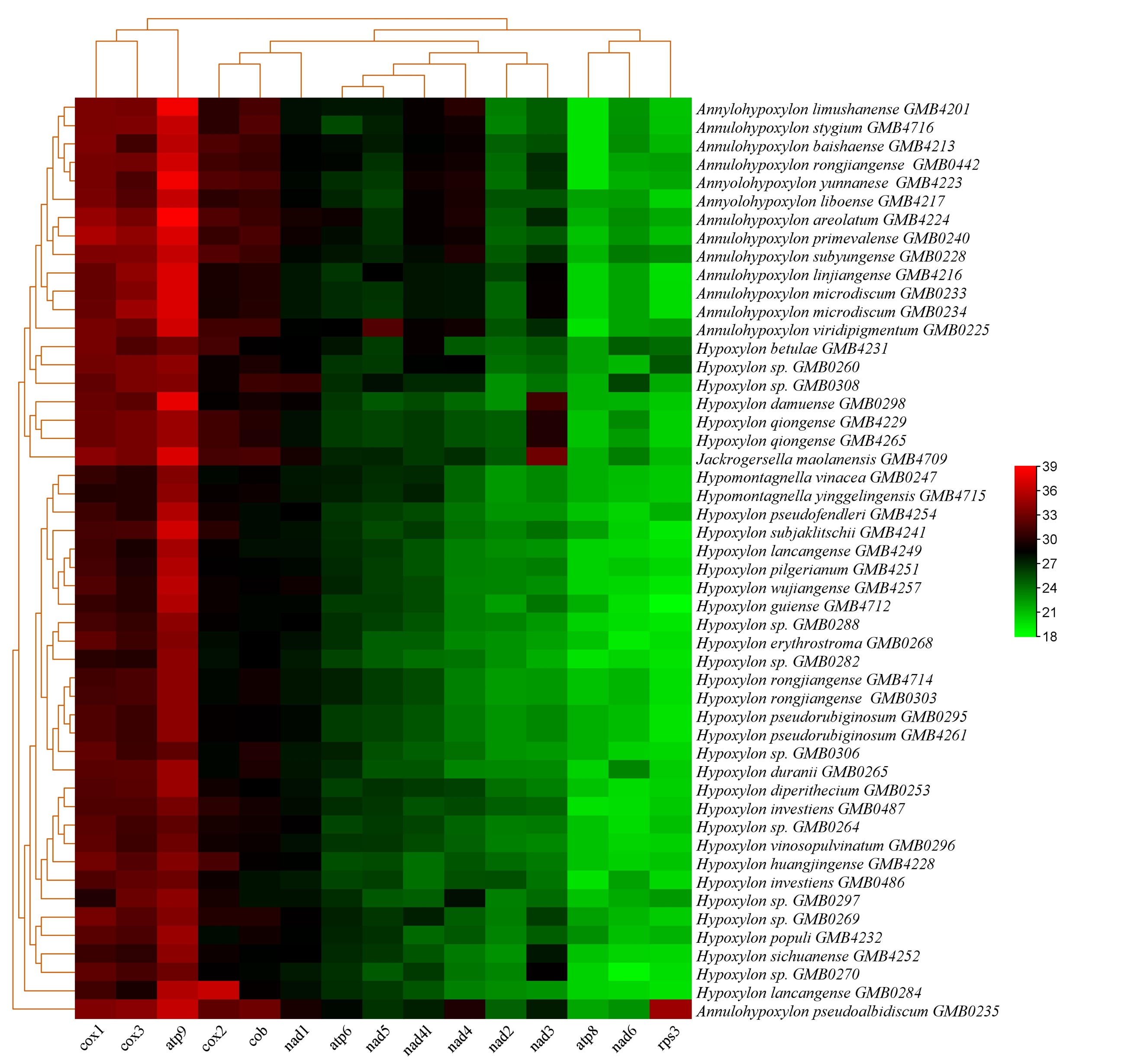
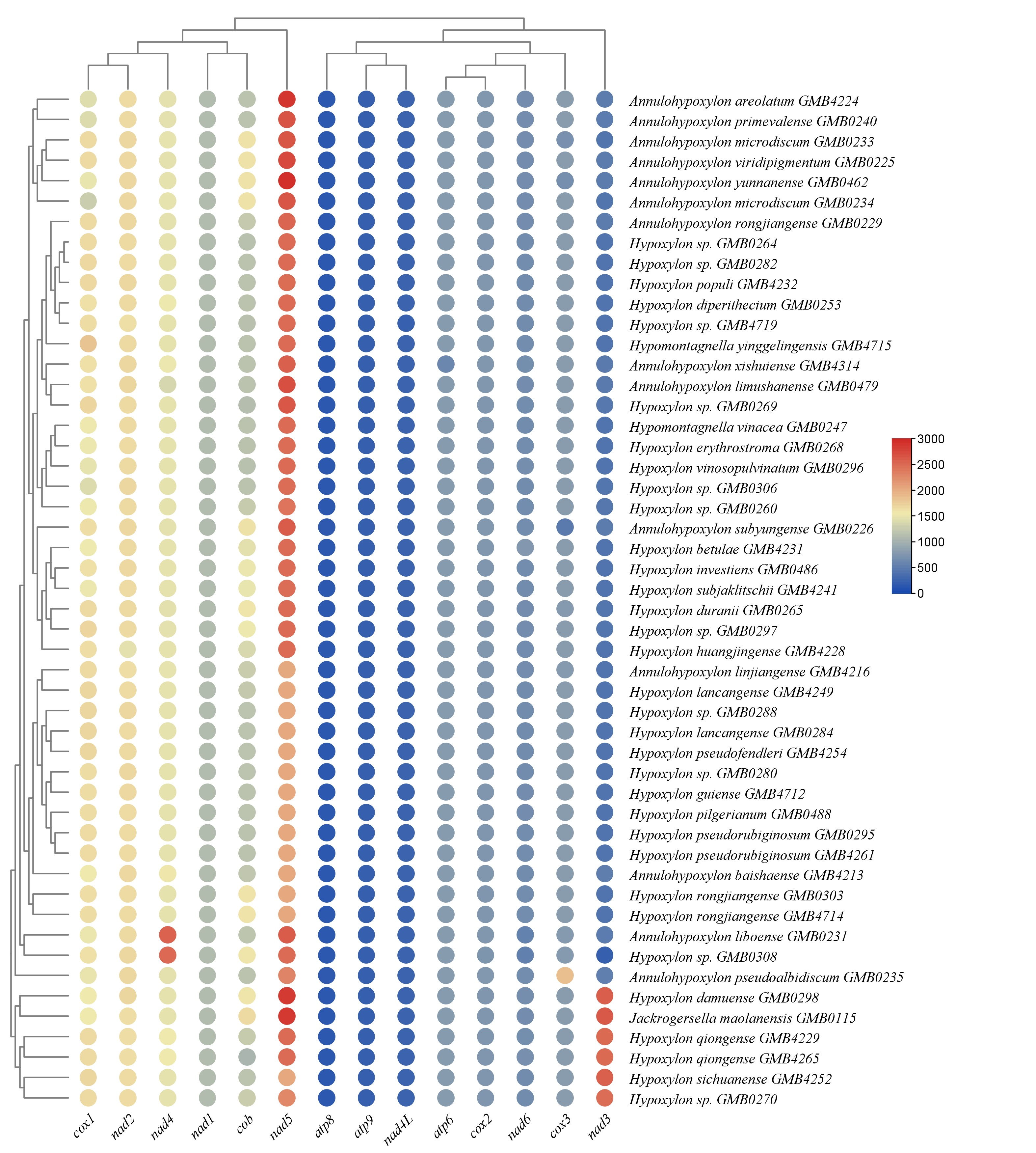



The structural patterns correlate with phylogenetic clades (Fig. 2). Clades M1, M2, and M3, which are predominantly composed of Annulohypoxylon species, contain the structurally complex mitogenomes. For example, A. viridipigmentum GMB0225 (Clade M1) has the highest number of homologous regions (39), suggesting an extensive history of break-and-rejoin events. In contrast, A. baishaense GMB4213 (Clade M2) shows reduced fragmentation (35 regions), indicating a possible shift toward structural stabilization in this lineage.
The genome size of Jackrogersella maolanensis GMB0115 (Fig. 2; Clade M4) is significantly larger than many species in Hypoxylon or Hypomontagnella. Structurally, Jackrogersella appears more similar to Annulohypoxylon than to its more immediate phylogenetic neighbors in the Hypoxylon complex. This high degree of fragmentation suggests that Jackrogersella represents a long-independent lineage where mitochondrial rearrangements have accumulated at a high rate, justifying its distinct classification at the genus level.
Species of Hypomontagnella, clustered in Clade M10 (Fig. 2), demonstrate a high degree of internal consistency. Hypomontagnella yinggelingensis GMB4715 (12 regions) and H. vinacea GMB0247 (11 regions) share nearly identical structural profiles. The slight variation in size and region count suggests a relatively recent divergence with minimal large-scale rearrangements. This stability in Clade M10 contrasts sharply with the volatility seen in the Annulohypoxylon clades (M1-M3), highlighting how different genera within the same family can follow distinct structural rules.
tRNA Genes in Hypoxylaceae Mitogenomes
The number of tRNA genes varied across the genera in the 50 Hypoxylaceae mitochondrial genomes. In the genus Annulohypoxylon, the number of tRNA genes ranged from 25 to 27. In the genus Hypomontagnella, 25 and 25 tRNA genes were detected in Hy. vinacea GMB0247 and Hy. yinggelingensis GMB4715 respectively. In the genus Hypoxylon, the number of tRNA genes ranged from 25 to 35. Hypoxylon sp. GMB0297 exhibited the highest number of tRNA genes (35), whereas H. guiense GMB4712, Hypoxylon sp. GMB0282, and Hypoxylon sp. GMB0306 each exhibited the lowest number, with 25 tRNA genes. In the genus Jackrogersella, J. maolanensis GMB0115 exhibited 25 tRNA genes. All tRNAs detected in the Hypoxylaceae mitogenomes were folded into classical cloverleaf structures (Supplementary Fig. 6). A representative example of the predicted tRNA secondary structure of Annulohypoxylon areolatum GMB4224 is shown in Fig. 28. The extra stem or loop regions were identified as the primary contributors to size variation among tRNA genes in Hypoxylaceae, with most variation occurring in the D-arm.
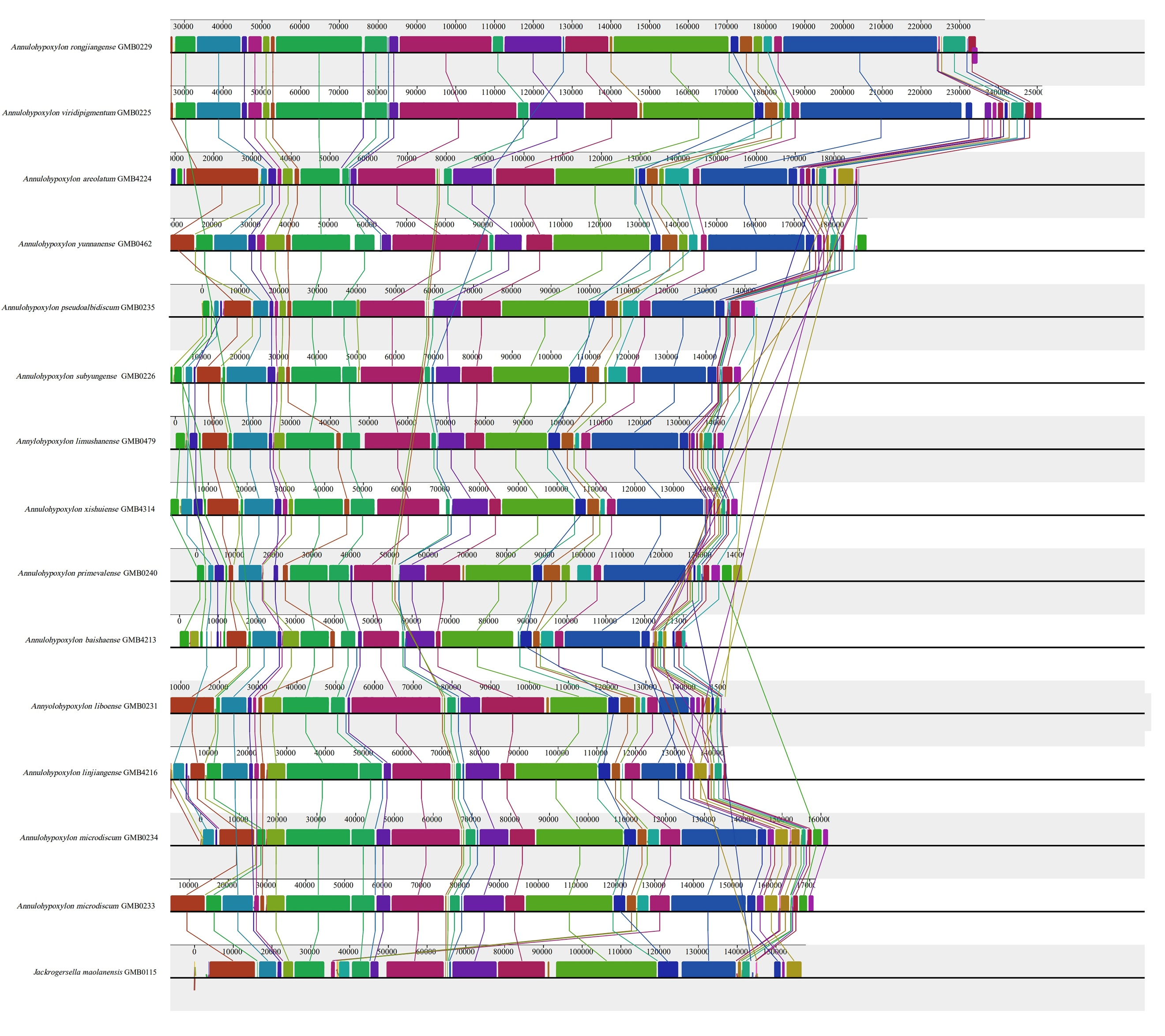
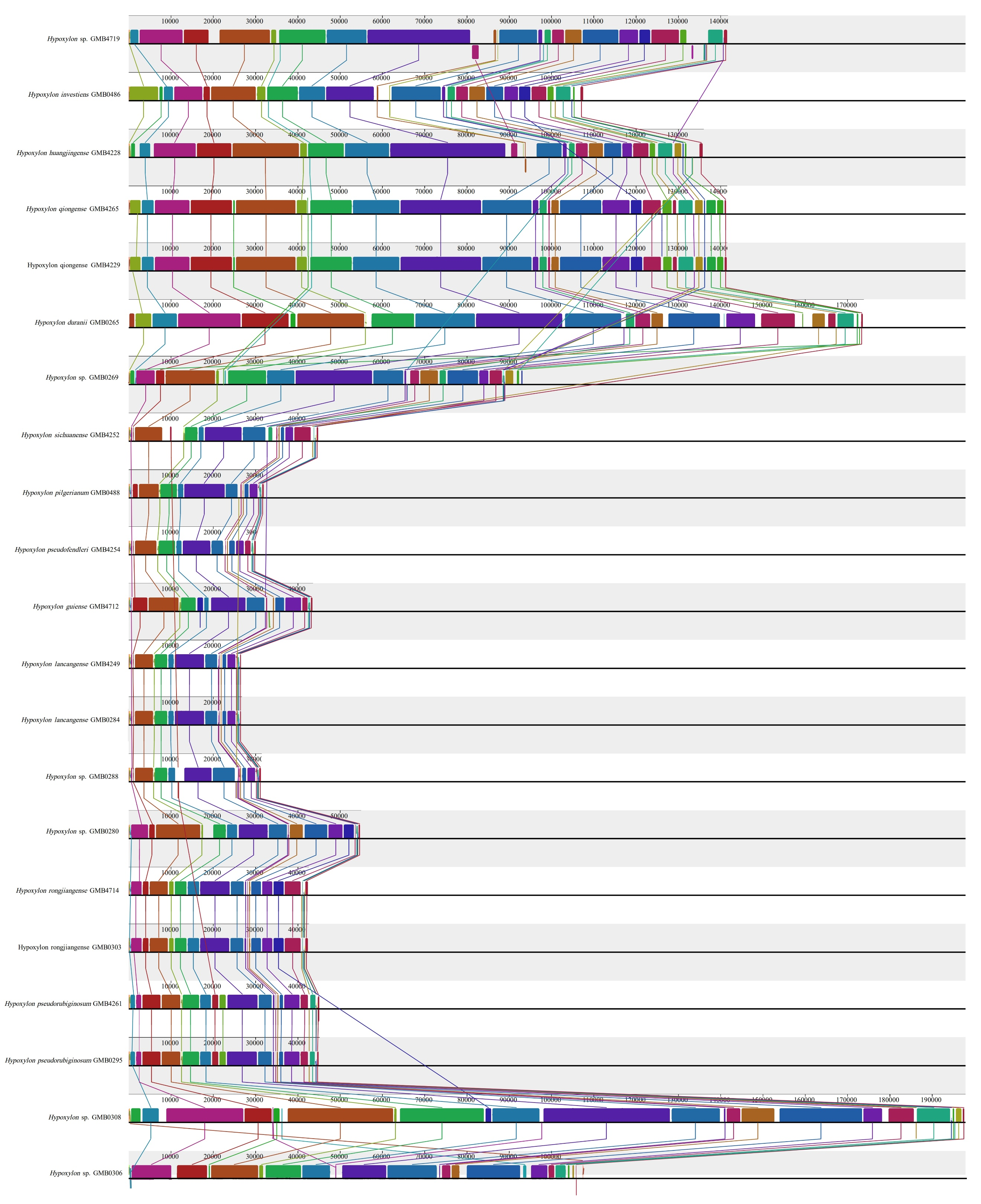
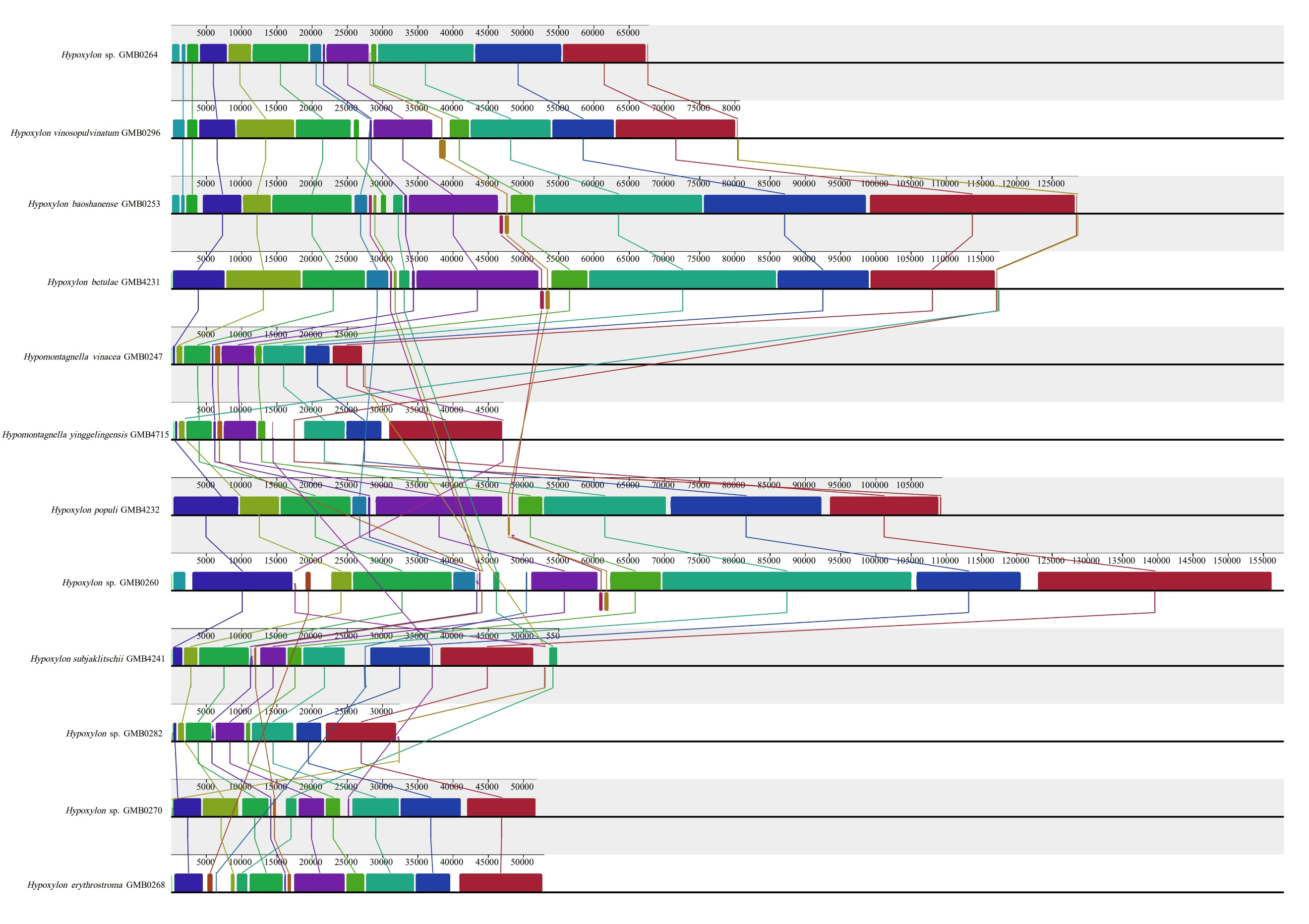
Discussion
The field of fungal taxonomy is dynamic and has been significantly impacted by technological and scientific advancements. In the seventeenth century, the invention of the microscope revolutionized the study of fungi by enabling researchers to observe previously inaccessible micromorphological features. By the twentieth century, physiological and biochemical assays such as color reactions and thin-layer chromatography (TLC) of secondary metabolites further refined species delimitations (Hsieh et al. 2005; Nilsson et al. 2008; Schoch et al. 2012; Houbraken et al. 2014).
The rise of molecular methods in the late twentieth century introduced DNA barcoding and phylogenetic reconstruction, which significantly improved taxonomic resolution. Early studies relied on single-locus datasets, typically ribosomal markers (18S rDNA, ITS1/2, 5.8S rDNA, 28S rDNA), but more recent multi-locus analyses including β-tubulin, translation elongation factor 1-α, and RNA polymerase II subunit rpb2 have yielded more robust and stable tree topologies (Dornburg et al. 2017; Zhang et al. 2017; Hyde et al. 2017; Wendt et al. 2018; Li et al. 2023, 2024). Together, these combined morphological, biochemical, and molecular approaches continue to refine classification of Hypoxylaceae and reveal cryptic diversity within the family.
Despite these advancements, the classification of Hypoxylaceae remains with challenges. Morphologically cryptic species often show subtle or overlapping phenotypic characters, while significant molecular variations are observed in commonly used markers such as ITS, LSU, rpb2, and tub2. Stadler et al. (2020), Wibberg et al. (2021), and Cedeño-Sanchez et al. (2024a) argue that the universally recognized ITS region, despite being the most used marker in fungal classification, has notable limitations in this group due to high intragenomic polymorphism and deep paralogy, which lead to ambiguous sequence alignments and conflicting phylogenetic interpretations. Consequently, Stadler and colleagues attempted to utilize the chemical composition of stroma as a basis for classification and published many articles (Lambert et al. 2019, Becker et al. 2020, Kuhnert et al. 2021, Cedeño-Sanchez et al. 2023). However, chemical classification has its own constraints, including high costs of chemical analysis equipment, more complex data analysis, the need for chemical experts’ assistance in compound structure analysis, unavailability of standard compounds, and difficulties in international specimen transfer. These challenges make it difficult to adopt chemical profiling as a universal method for fungal classification.
This study proposes mitochondrial genome (mitogenome) data as a supplementary criterion for species delimitation in Hypoxylaceae, integrating it with traditional morphological and multi-locus molecular data. Mitogenomes offer distinct advantages, including high copy numbers, structural variability and conserved protein-coding regions that are less prone to homoplasy compared to nuclear markers. Our comprehensive analysis of 50 Hypoxylaceae mitogenomes revealed extraordinary variability in genome size (26,809–251,279 bp), intron content (0–68 introns), intergenic region length, gene arrangement, and tRNA composition, highlighting pronounced structural plasticity across the family. This structural diversity, including genus- and species-specific patterns in intron content, gene order, and tRNA composition, provides a rich source of comparative characters.
Several species of Hypoxylon have been provisionally identified using the term "cf." (confer), reflecting their ambiguous but close relationship to known species. For example, taxa closely resembling H. ticinense share nearly identical teleomorph, and many specimens previously assigned to H. rubiginosum (often reported as H. cf. rubiginosum) show clear genetic divergence yet lack consistent morphological or chemical differences (Pourmoghaddam et al. 2020). Our results demonstrate that mitogenomic features particularly intron profiles, genome size, and intergenic region expansion provide additional discriminatory power for such cryptic taxa.
To differentiate such cryptic or closely related taxa, mitochondrial genomes can serve as supplementary barcodes, providing additional resolution beyond nuclear markers and morphology. As shown in Supplementary Tables 10 and 11, mitogenomic data reveal significant variation in genome size, intron content, and intergenic region lengths. Genome size variation was driven primarily by non-coding regions rather than protein-coding genes, as coding sequence lengths remained relatively conserved (CV 0–15%) compared to total genome size (CV ~50%). This genomic divergence supports the utility of mitogenomes in resolving species boundaries within Hypoxylon. Specifically, the extreme variability in genes like nad3 (225–2,631 bp) and the presence of unique, conserved gene rearrangements (e.g., the cox2 translocation in Jackrogersella maolanensis) offer powerful synapomorphies for clade and species discrimination. To distinguish these cryptic or closely allied taxa, mitogenomic data provide a promising solution. As demonstrated in Supplementary Table 10 and 11 and Supplementary Tables 1–7, significant variation in genome size, intron content, and intergenic region length was observed across different species. These differences support the potential of mitogenomes as supplementary barcodes to enhance taxonomic resolution.
Phylogenetic trees constructed using both nuclear multi-gene datasets (ITS-LSU-rpb2-tub2) and mitogenomic data showed largely consistent topologies, except for the taxonomic placement of a few individual species. For example, Hypomontagnella forms a monophyletic clade in nuclear gene-based phylogenies (Fig. 1) but appears as a well-supported sister lineage to Hypoxylon populi in mitogenomic analyses (Fig. 2). This discrepancy may result from the limited availability of mitogenomic data for many taxa, underscoring the need for broader sampling to resolve conflicting evolutionary signals and refine taxonomic placements.
Comparative analysis of mitochondrial gene sequences revealed substantial intra- and inter-specific length polymorphisms, particularly in intron-rich regions, alongside conserved protein-coding domains. Genes such as atp9, nad1, nad4L, and atp8 were highly conserved in length and showed low Ka/Ks ratios, indicating strong purifying selection, whereas genes nad3, nad5, cox1, and cob showed elevated variability and signatures of positive selection in certain lineages. These variations provide valuable information that can improve species delimitation, clarify evolutionary relationships, and enhance the resolution of taxonomic classifications within closely related or cryptic lineages. The case of the two Annulohypoxylon microdiscum strains (GMB0233 & GMB0234), which are morphologically identical yet show mitogenomic divergence, powerfully illustrates the potential of mitogenomes to reveal hidden genetic divergence and flag potential cryptic species. Future studies should prioritize the generation of mitogenomes for underrepresented taxa, enabling the development of a unified taxonomic framework that synergizes genomic, morphological, and chemical data. Establishing a standardized pipeline for mitogenome annotation and comparative analysis will be crucial for its adoption as a routine tool in fungal taxonomy. This article presents preliminary mitogenomic data, and the in-depth comparative analysis will be published in subsequent articles.
Conclusion
This study advances the taxonomy of Hypoxylaceae by using mitochondrial genome data with traditional morphological and molecular approaches. Through comprehensive analyses of 50 specimens, we identified 16 novel species and generated the first mitogenomes for Jackrogersella, Hypomontagnella, and Hypoxylon. This represents the most extensive mitogenomic dataset for Hypoxylaceae to date. Our analyses revealed remarkable mitogenomic diversity in genome size, gene order, and intron content, and the 14-core protein-coding genes may serve as valuable supplementary barcodes. These mitogenomic variations, alongside collinearity and sequence divergence, provide robust support for distinguishing closely related taxa. Genes such as nad3, nad5, cox1, and cob, which show elevated variability and signatures of positive selection, emerge as promising supplementary barcode candidates. A more detailed analysis of these genes will be provided in a forthcoming publication. While nuclear multi-locus phylogenies and mitogenomic trees largely aligned, discrepancies in clade placements (e.g., Hypomontagnella) highlight the impact of limited mitogenome sampling and underscore the need for broader taxonomic representation across the family. The extensive structural diversity observed from compact, intron-less genomes in some Hypoxylon species to massive, intron-rich genomes in Annulohypoxylon demonstrates that mitogenome itself contains phylogenetically informative signals. To build a unified and stable classification for Hypoxylaceae, future work should prioritize expanding mitogenome sequencing alongside detailed morphological and chemical profiling across the family. Such an integrative framework will be essential for resolving cryptic species complexes, understanding mitochondrial evolution, and developing reliable mitochondrial barcode systems for fungal taxonomy.
Acknowledgements
This research was supported by the National Natural Science Foundation of China (32170019 32460007, 31960005, and 32000009); the Funded Research Projects of the State Key Laboratory of Discovery and Utilization of Functional Components in Traditional Chinese Medicine 2025 (GMUSKL-202510); the Guizhou Medical University High-Level Talent Launch Fund Project (2023-058); the Guizhou Provincial Scientific and Technologic Innovation Base (No. [2023]003); the High-level Innovation Talents of Guizhou (No. GCC [2023]048); National Natural Science Foundation of China (12132006); the Guizhou Provincial Natural Science Foundation for High-Level Innovative Talents and Teams (2016-5676, 2015-4021).
Author contributions
All authors contributed to the study conception and design. Conceptualization: J.-C. Kang, Q.-R. Li, Z.-B. Meng. Collection and morphological examinations: W.-Y. Zen, X. Zhou, Y.-L. Ren, Y.-H. Pi, H.-M. Hu, L.-L. Liu. Molecular sequencing, and phylogenetic analyses: X. Zhou, H.-M. Hu, K. Habib. Specimen identification: K. Habib, Q.-R. Li. Original draft preparation: K. Habib, Q.-R. Li. Review, supervision: Y.-Q. Kang, J.-C. Kang, X.-C. Shen. Editing: K. Habib, K.D. Hyde. All authors have read and agreed to the published version of the manuscript.
ORCID
Wen-Yu Zeng: https://orcid.org/0009-0001-6826-7591
Xin Zhou: https://orcid.org/0009-0002-4204-1225
Kamran Habib: https://orcid.org/0000-0003-2572-0306
Ze-Bin Meng: https://orcid.org/0000-0002-9995-6605
Qi-Rui Li: https://orcid.org/0000-0001-8735-2890
Ji-Chuan Kang: https://orcid.org/0000-0002-6294-5793
Conflict of interest statement
The author list includes members of the Editorial Board of Fungal Diversity. They were not involved in the journal’s review of, or decisions related to, this manuscript. The authors declare no competing interests.
Data availability
The data sets generated during and/or analyzed during the current study are available in the MycoBank repository (included in the manuscript), and GenBank (included in Supplementary Table 9). And also, the data sets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Supplemental information
The online version contains supplemental information available at https://doi.org/10.65390/fdiv.2026.136011.
Rights and permissions
The Author(s) 2026. Published by BioAcademic Press on behalf of Kunming Institute of Botany,Chinese Academy of Sciences (CAS) and Mushroom Research Foundation. This is an open accessarticle under the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0), which permits use, distribution and reproduction in any medium, provided the originalwork is properly cited.
References
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410. https://doi.org/10.1016/S0022-2836(05)80360-2
- Becker K, Wessel AC, Luangsa-Ard JJ, Stadler M (2020) Viridistratins A–C, antimicrobial and cytotoxic benzo[j]fluoranthenes from stromata of Annulohypoxylon viridistratum (Hypoxylaceae, Ascomycota). Biomolecules 10:805. https://doi.org/10.3390/biom10050805
- Bernt M, Donath A, Jühling F, Externbrink F, Florentz C, Fritzsch G et al (2013) MITOS: improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol 69:313–319. https://doi.org/10.1016/j.ympev.2012.08.023
- Bills GF, González-Menéndez V, Martín J, Platas G, Fournier J, Peršoh D, Stadler M (2012) Hypoxylon pulicicidum sp. nov. (Ascomycota, Xylariales), a pantropical insecticide-producing endophyte. PLoS ONE 7:e46687. https://doi.org/10.1371/journal.pone.0046687
- Caspermeyer J (2016) MEGA evolutionary software re-engineered to handle today's big data demands. Mol Biol Evol 33(7):1887. https://doi.org/10.1093/molbev/msw074
- Cedeño-Sanchez M, Charria-Girón E, Lambert C, Luangsa-Ard JJ, Decock C, Franke R et al (2023) Segregation of the genus Parahypoxylon (Hypoxylaceae, Xylariales) from Hypoxylon by a polyphasic taxonomic approach. MycoKeys 95:131–162. https://doi.org/10.3897/mycokeys.95.98125
- Cedeño-Sanchez M, Cheng T, Lambert C, Kolarík M, Kuhnert E, Cox RJ et al (2024a) Unraveling intragenomic polymorphisms in the high-quality genome of Hypoxylaceae: a comprehensive study of the rDNA cistron. Mycol Prog 23:5. https://doi.org/10.1007/s11557-023-01940-2
- Cedeño-Sanchez M, Lambert C, Mejia LC, Ebada SS, Stadler M (2024b) Chemotaxonomic and molecular phylogenetic studies of selected Hypoxylon species from the Neotropics. Mycology 16:250–265. https://doi.org/10.1080/21501203.2024.2378071
- Chen TQ, Yang C, Xu XL, Yang L, He HQ, Weng MT et al (2024) Comparative mitogenomics provides valuable insights for the phylogeny and new DNA barcodes of Ganoderma. J Fungi 10(11):769. https://doi.org/10.3390/jof10110769
- Cheng J, Luo Q, Ren Y, Luo Z, Liao W, Wang X et al (2021) Panorama of intron dynamics and gene rearrangements in the phylum Basidiomycota as revealed by the complete mitochondrial genome of Turbinellus floccosus. Appl Microbiol Biotechnol 105(5):2017–2032. https://doi.org/10.1007/s00253-021-11153-w
- Cruz KDS, Lima MC, de Jesus MA, de Souza AQ, Sales-Campos C (2021) Annulohypoxylon (Hypoxylaceae, Ascomycota) from Amazonian-forest of Brazil, with a description of one new species. N Z J Bot 59:267–284. https://doi.org/10.1080/0028825X.2020.1846570
- Dai DQ, Phookamsak R, Wijayawardene NN, Li WJ, Bhat DJ, Xu JC et al (2017) Bambusicolous fungi. Fungal Divers 82:1–105. https://doi.org/10.1007/s13225-016-0367-8
- Darling AE, Mau B, Perna NT (2010) Progressive Mauve: multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 5:e11147. https://doi.org/10.1371/journal.pone.0011147
- Dayarathne MC, Jones EBG, Maharachchikumbura SSN, Devadatha B, Sarma VV, Khongphinitbunjong K et al (2020) Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 11:1–188. https://doi.org/10.5943/mycosphere/11/1/1
- Dornburg A, Townsend JP, Wang Z (2017) Maximizing power in phylogenetics and phylogenomics: a perspective illuminated by fungal big data. Adv Genet 100:1–47. https://doi.org/10.1016/bs.adgen.2017.09.007
- Férandon C, Moukha S, Callac P, Benedetto JP, Castroviejo M, Barroso G (2010) The Agaricus bisporus cox1 gene: the longest mitochondrial gene and the largest reservoir of mitochondrial group I introns. PLoS ONE 5(11):e14048. https://doi.org/10.1371/journal.pone.0014048
- Fonseca PL, Badotti F, De-Paula RB, Araújo DS, Bortolini DE, Del-Bem LE et al (2020) Exploring the relationship among divergence time and coding and non-coding elements in the shaping of fungal mitochondrial genomes. Front Microbiol 11:765. https://doi.org/10.3389/fmicb.2020.00765
- Fournier J, Lechat C, Pelissier M (2014) Hypoxylon flavocremeum sp. nov (Xylariaceae), a new species from Mayotte (France outre mer), with notes on some other Hypoxylon spp. collected in this island. Ascomycete. org. 6(4):65-71. https://doi.org/10.25664/art-0102
- Fournier J, Köpcke B, Stadler M (2010) New species of Hypoxylon from western Europe and Ethiopia. Mycotaxon 113:209–235. https://doi.org/10.5248/113.209
- Fournier J, Lechat C (2016) Some Annulohypoxylon spp. (Xylariaceae) from French Guiana, including three new species. Ascomycete.org 8:33–53. https://doi.org/10.25664/ART-0169
- Fournier J, Lechat C, Courtecuisse R (2015) The genus Hypoxylon (Xylariaceae) in Guadeloupe and Martinique (French West Indies). Ascomycete. org, 7(5) 145 212. https://doi.org/10.25664/art-0140
- Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes—application to the identification of mycorrhizae and rusts. Mol Ecol 2:113–118.
- Glass NL, Donaldson GC (1995) Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl Environ Microbiol 61:1323–1330.
- Grant JR, Enns E, Marinier E, Mandal A, Herman EK, Chen C et al (2023) Proksee: in-depth characterization and visualization of bacterial genomes. Nucleic Acids Res 51(W1):W484–W492. https://doi.org/10.1093/nar/gkad326
- Hall TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41:95–98.
- Houbraken J, de Vries RP, Samson RA (2014) Modern taxonomy of biotechnologically important Aspergillus and Penicillium species. Adv Appl Microbiol 86:199–249. https://doi.org/10.1016/B978-0-12-800262-9.00004-4
- Hsieh HM, Ju YM, Rogers JD (2005) Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 97:844–865. https://doi.org/10.1080/15572536.2006.11832776
- Hyde KD, Norphanphoun C, Abreu VP, Bazzicalupo A, Chethana KT, Clericuzio M et al (2017) Fungal diversity notes 603–708: taxonomic and phylogenetic notes on genera and species. Fungal Divers 87:1–235. https://doi.org/10.1007/s13225-017-0391-3
- Ju YM, Rogers JD (1996) A revision of the genus Hypoxylon. Mycologia Memoir No.20. APS Press, St. Paul.
- Katoh K, Rozewicki J, Yamada KD (2019) MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform 20:1160–1166. https://doi.org/10.1093/bib/bbx108
- Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S et al (2012) Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28:1647–1649. https://doi.org/10.1093/bioinformatics/bts199
- Kryazhimskiy S, Plotkin JB (2008) The population genetics of dN/dS. PLoS Genet 4:e1000304. https://doi.org/10.1371/journal.pgen.1000304
- Kuhnert E, Fournier J, Peršoh D, Luangsa-Ard JJD, Stadler M (2014) New Hypoxylon species from Martinique and new evidence on the molecular phylogeny of Hypoxylon based on ITS rDNA and β-tubulin data. Fungal Divers 64:181–203. https://doi.org/10.1007/s13225-013-0264-3
- Kuhnert E, Surup F, Sir EB, Lambert C, Hyde KD, Hladki AI et al (2015) Lenormandins A–G, new azaphilones from Hypoxylon lenormandii and Hypoxylon jaklitschii sp. nov., recognised by chemotaxonomic data. Fungal Divers 71:165–184. https://doi.org/10.1007/s13225-014-0318-1
- Kuhnert E, Navarro-Muñoz JC, Becker K, Stadler M, Collemare J, Cox RJ (2021) Secondary metabolite biosynthetic diversity in the fungal family Hypoxylaceae and Xylaria hypoxylon. Stud Mycol 99:100118. https://doi.org/10.1016/j.simyco.2021.100118
- Lambert C, Pourmoghaddam MJ, Cedeño-Sanchez M, Surup F, Khodaparast SA, Krisai-Greilhuber I et al (2021) Resolution of the Hypoxylon fuscum complex (Hypoxylaceae, Xylariales) and discovery and biological characterization of two of its prominent secondary metabolites. J Fungi 7:131. https://doi.org/10.3390/jof7020131
- Lambert C, Wendt L, Hladki AI, Stadler M, Sir EB (2019) Hypomontagnella (Hypoxylaceae): a new genus segregated from Hypoxylon by a polyphasic taxonomic approach. Mycol Prog 18:187–201. https://doi.org/10.1007/s11557-018-1452-z
- Lanfear R, Frandsen PB, Wright AM, Senfeld T, Calcott B (2017) PartitionFinder 2: new methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol Biol Evol 34 (3):772-773. https://doi.org/10.1093/molbev/msw260
- Leman-Loubière C, Le Goff G, Debitus C, Ouazzani J (2017) Sporochartines A–E, a new family of natural products from the marine fungus Hypoxylon monticulosum isolated from a Sphaerocladina sponge. Front Mar Sci 4:399. https://doi.org/10.3389/fmars.2017.00399
- Li Q, Xiang D, Wan Y, Wu Q, Wu X, Ma C et al (2019) The complete mitochondrial genomes of five important medicinal Ganoderma species: features, evolution, and phylogeny. Int J Biol Macromol 139:397–408. https://doi.org/10.1016/j.ijbiomac.2019.08.003
- Li Q, Ren Y, Xiang D, Shi X, Zhao J, Peng L, Zhao G (2020) Comparative mitogenome analysis of two ectomycorrhizal fungi (Paxillus) reveals gene rearrangement, intron dynamics, and phylogeny of basidiomycetes. IMA Fungus 11(1):12. https://doi.org/10.1186/s43008-020-00038-8.
- Li QR, Pi YH, Charria-Girón E, Habib K, Hu HM, Wongkanoun S, Stadler M, Liu LL, Shen XC, Kang JC (2025). Polyphasic taxonomy and chemical diversity of Annulohypoxylon (Ascomycota, Hypoxylaceae): New species from China’s tropical forest. Persoonia 55: 159–201. https://doi.org/10.3114/persoonia.2025.55.05
- Li QR, Long SH, Lin Y, Wu YP, Wu QZ, Hu HM, Shen XC, Zhang X, Wijayawardene NN, Kang JC (2023) Diversity, morphology, and molecular phylogeny of Diatrypaceae from southern China. Front Microbiol 14:1140190. https://doi.org/10.3389/fmicb.2023.1140190
- Li QR, Habib K, Long SH, Wu YP, Zhang X, Hu HM et al (2024) Unveiling fungal diversity in China: new species and records within the Xylariaceae. Mycosphere 15:275–364. https://doi.org/10.5943/mycosphere/15/1/2
- Liu YJ, Whelen S, Hall BD (1999) Phylogenetic relationships among ascomycetes: evidence from an RNA polymerase II subunit. Mol Biol Evol 16:1799–1808.
- Lowe TM, Chan PP (2016) tRNAscan-SE On-line: integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res 44(W1):W54–W57. https://doi.org/10.1093/nar/gkw413
- Ma H, Song Z, Pan X, Li Y, Yang Z, Qu Z (2022) Multi-gene phylogeny and taxonomy of Hypoxylon (Hypoxylaceae, Ascomycota) from China. Diversity 14:37. https://doi.org/10.3390/d14010037
- Medina RP, Silva AD, Andersen RJ, Araújo AR, Silva DHS (2016) Botryane sesquiterpenes and binaphthalene tetrols from endophytic fungi associated to the marine red algae Asparagopsis taxiformis. Planta Med 82:P562. https://doi.org/10.1055/s-0036-1596629
- Miller JH (1961) A monograph of the world species of Hypoxylon. Univ. Georgia Press, Athens. https://archive.org/details/monographofworld0000juli
- Miller MA, Pfeiffer W, Schwartz T (2010) Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In: 2010 Gateway Computing Environments Workshop (GCE). IEEE, pp 1–8. https://doi.org/10.1109/GCE.2010.5676129
- Mühlbauer A, Triebel D, Persoh D, Wollweber H, Seip S, Stadler M (2002) Macrocarpones, novel metabolites from stromata of Hypoxylon macrocarpum, and new evidence on the chemotaxonomy of Hypoxylon species. Mycol Prog 1:235–248. https://doi.org/10.1007/s11557-006-0021-z
- Nilsson RH, Kristiansson E, Ryberg M, Hallenberg N, Larsson KH (2008) Intraspecific ITS variability in the kingdom Fungi as expressed in the international sequence databases and its implications for molecular species identification. Evol Bioinform 4:EBO–S653. https://doi.org/10.4137/EBO.S653
- O’Donnell K, Cigelnik E (1997) Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol Phylogenet Evol 7:103–116 https://doi.org/10.1006/mpev.1996.0376
- Pažoutová S, Follert S, Bitzer J, Keck M, Surup F, Šrůtka P et al (2013) A new endophytic insect-associated Daldinia species, recognized from a comparison of secondary metabolite profiles and molecular phylogeny. Fungal Divers 60:107–123. https://doi.org/10.1007/s13225-013-0238-5
- Phookamsak R, Hyde KD, Jeewon R, Bhat DJ, Jones EG, Maharachchikumbura SS et al (2019) Fungal diversity notes 929–1035: taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers 95:1–273. https://doi.org/10.1007/s13225-019-00421-w
- Pourmoghaddam MJ, Lambert C, Surup F, Khodaparast SA, Krisai-Greilhuber I, Voglmayr H et al (2020) Discovery of a new species of the Hypoxylon rubiginosum complex from Iran and antagonistic activities of Hypoxylon spp. against the Ash Dieback pathogen, Hymenoscyphus fraxineus, in dual culture. MycoKeys 66:105–133. https://doi.org/10.3897/mycokeys.66.50946
- Rambaut A (2012) FigTree: Tree Figure drawing tool 2006–2012, version 1.4.0. Institute of Evolutionary Biology, University of Edinburgh, Edinburgh.
- Rayner RW (1970). A mycological colour chart. Kew and British Mycological Society, Commonwealth Mycological Institute, Kew. https://www.cabidigitallibrary.org https://doi/full/10.5555/19711606619
- Rogers JD (1981) Two new Hypoxylon species from Gabon. Can J Bot 59:1363–1364.
- Ronquist F, Teslenko M, Van Der Mark P, Ayres DL, Darling A, Höhna S et al (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol 61:539–542. https://doi.org/10.1093/sysbio/sys029
- Rozas J, Ferrer-Mata A, Sánchez-DelBarrio JC, Guirao-Rico S, Librado P, Ramos-Onsins SE et al (2017) DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol Biol Evol 34:3299–3302. https://doi.org/10.1093/molbev/msx248
- Samarakoon MC, Hyde KD, Maharachchikumbura SS, Stadler M, Jones EBG, Promputtha I et al (2022) Taxonomy, phylogeny, molecular dating and ancestral state reconstruction of Xylariomycetidae (Sordariomycetes). Fungal Divers 112:1–88. https://doi.org/10.1007/s13225-021-00495-5
- Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, Levesque CA et al (2012) Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi. Proc Natl Acad Sci USA 109:6241–6246. https://doi.org/10.1073/pnas.1117018109
- Senanayake IC, Rathnayaka AR, Marasinghe DS, Calabon MS, Gentekaki E, Lee HB et al (2020) Morphological approaches in studying fungi: collection, examination, isolation, sporulation and preservation. Mycosphere 11:2678–2754. https://doi.org/10.5943/mycosphere/11/1/20
- Sir EB, Kuhnert E, Hladki AI, Romero AI (2018) Annulohypoxylon (Hypoxylaceae) species from Argentina. Darwiniana 6:68–83. https://doi.org/10.14522/darwiniana.2018
- Sir EB, Becker K, Lambert C, Bills GF, Kuhnert E (2019) Observations on Texas hypoxylons, including two new Hypoxylon species and widespread environmental isolates of the H. croceum complex identified by a polyphasic approach. Mycologia 111:832–856. https://doi.org/10.1080/00275514.2019.1637705
- Smith DR (2016) The past, present and future of mitochondrial genomics: have we sequenced enough mtDNAs? Brief Funct Genomics 15:47–54. https://doi.org/10.1093/bfgp/elv027
- Song ZK, Zhu AH, Liu ZD, Qu Z, Li Y, Ma HX (2022) Three new species of Hypoxylon (Xylariales, Ascomycota) on a multigene phylogeny from Medog in Southwest China. J Fungi 8:500. https://doi.org/10.3390/jof8050500
- Stadler M, Wollweber H, Fournier J (2004) A host-specific species of Hypoxylon from France, and notes on the chemotaxonomy of the “Hypoxylon rubiginosum complex”. Mycotaxon 90:187–211.
- Stadler M, Lambert C, Wibberg D, Kalinowski J, Cox RJ, Kolařík M, Kuhnert E (2020) Intragenomic polymorphisms in the ITS region of high-quality genomes of the Hypoxylaceae (Xylariales, Ascomycota). Mycol Prog 19:235–245. https://doi.org/10.1007/s11557-019-01552-9
- Stamatakis A (2015) Using RAxML to infer phylogenies. Curr Protoc Bioinform 51:6.14s. https://doi.org/10.1002/0471250953.bi0614s51
- Stothard P (2000) The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. Biotechniques 28(6):1102–1104. https://doi.org/10.2144/00286ir01
- Tamura K, Stecher G, Kumar S (2021) MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol 38:3022–3027. https://doi.org/10.1093/molbev/msab120
- Tan YP, Shivas RG (2022) Nomenclatural novelties. Index of Australian Fungi 1:1–18.
- Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22 (22):4673–4680. https://doi.org/10.1093/nar/22.22.4673
- U’Ren JM, Miadlikowska J, Zimmerman NB, Lutzoni F, Stajich JE, Arnold AE (2016) Contributions of north American endophytes to the phylogeny, ecology, and taxonomy of Xylariaceae (Sordariomycetes, Ascomycota). Mol Phylogenet Evol 98:210–232. https://doi.org/10.1016/j.ympev.2016.02.010
- Valach M, Burger G, Gray MW, Lang BF (2014) Widespread occurrence of organelle genome-encoded 5S rRNAs including permuted molecules. Nucleic Acids Res 42:13764–13777. https://doi.org/10.1093/nar/gku1266
- Vaidya G, Lohman DJ, Meier R (2011) SequenceMatrix: concatenation software for the fast assembly of multi‐gene datasets with character set and codon information. Cladistics 27(2):171–180. https://doi.org/10.1111/j.1096-0031.2010.00329.x
- Vilgalys R, Hester M (1990) Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J Bacteriol 172:4238–4246.
- Wang J, Zhang L, Zhang Q-L, Zhou MQ, Wang XT, Yang XZ, Yuan ML (2017) Comparative mitogenomic analysis of mirid bugs (Hemiptera: Miridae) and evaluation of potential DNA barcoding markers. PeerJ 5:e3661. https://doi.org/10.7717/peerj.3661
- Wendt L, Sir EB, Kuhnert E, Heitkämper S, Lambert C, Hladki AI et al (2018) Resurrection and emendation of the Hypoxylaceae, recognised from a multigene phylogeny of the Xylariales. Mycol Prog 17:115–154. https://doi.org/10.1007/s11557-017-1311-3
- White TJ, Bruns T, Lee SJWT, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR Protocols: a guide to methods and applications. Academic Press, San Diego, pp 315–322.
- Wibberg D, Stadler M, Lambert C, Bunk B, Spröer C, Rückert C et al (2021) High quality genome sequences of thirteen Hypoxylaceae (Ascomycota) strengthen the phylogenetic family backbone and enable the discovery of new taxa. Fungal Divers 106:7–28. https://doi.org/10.1007/s13225-020-00447-5
- Wu P, Yao T, Ren Y, Ye J, Qing Y, Li Q, Gui M (2021) Evolutionary insights into two widespread ectomycorrhizal fungi (Pisolithus) from comparative analysis of mitochondrial genomes. Front Microbiol 12:583129. https://doi.org/10.3389/fmicb.2021.583129
- Zhang N, Luo J, Bhattacharya D (2017) Advances in fungal phylogenomics and their impact on fungal systematics. Adv Genet 100:309–328. https://doi.org/10.1016/bs.adgen.2017.09.004